Wu Yongrui, Messing Joachim
Institute of Plant Physiology and Ecology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences Shanghai, China.
Waksman Institute of Microbiology, Rutgers University Piscataway, NJ, USA.
Front Plant Sci. 2014 May 30;5:240. doi: 10.3389/fpls.2014.00240. eCollection 2014.
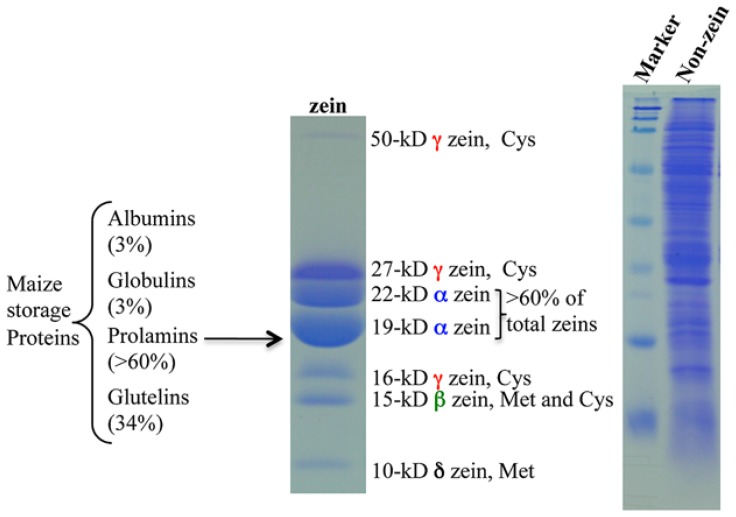
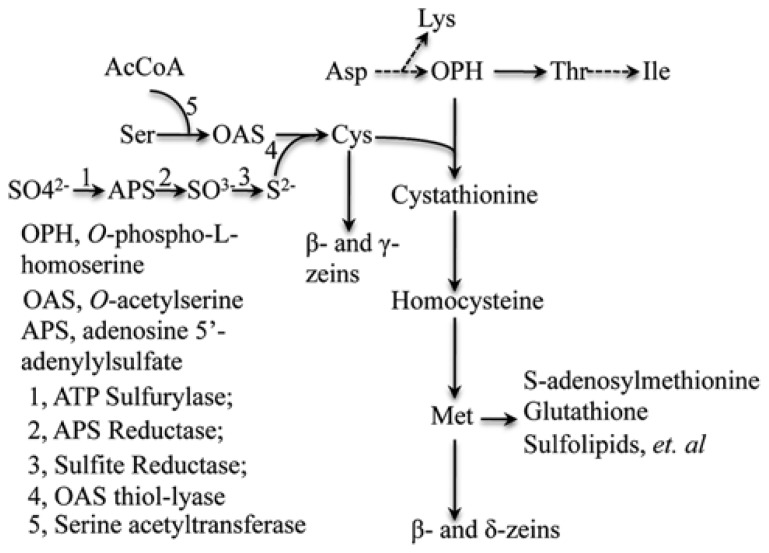
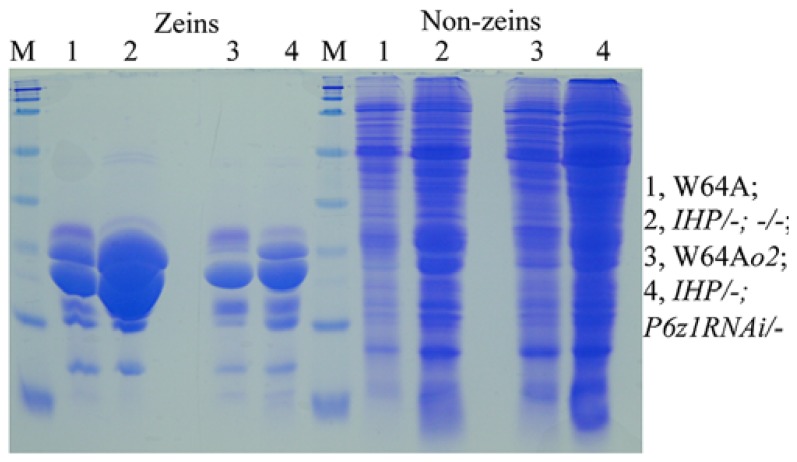
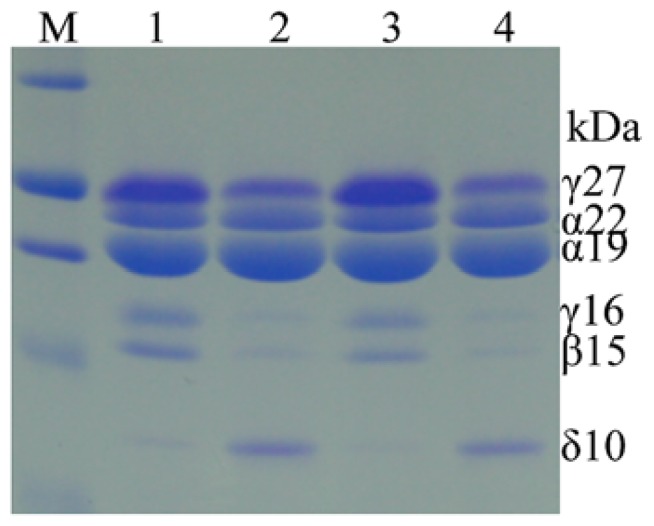
Most flowering plant seeds are composed of the embryo and endosperm, which are surrounded by maternal tissue, in particular the seed coat. Whereas the embryo is the dormant progeny, the endosperm is a terminal organ for storage of sugars and amino acids in proteins and carbohydrates, respectively. Produced in maternal leaves during photosynthesis, sugars, and amino acids are transported to developing seeds after flowering, and during germination they nourish early seedlings growth. Maize endosperm usually contains around 10% protein and 70% starch, and their composition ratio is rather stable, because it is strictly regulated through a pre-set genetic program that is woven by networks of many interacting or counteracting genes and pathways. Endosperm protein, however, is of low nutritional value due mainly to the high expression of the α-zein gene family, which encodes lysine-free proteins. Reduced levels of these proteins in the opaque 2 (o2) mutant and α-zein RNAi (RNA interference) transgenic seed is compensated by an increase of non-zein proteins, leading to the rebalancing of the nitrogen sink and producing more or less constant levels of total proteins in the seed. The same rebalancing of zeins and non-zeins has been observed for maize seeds bred for 30% protein. In contrast to the nitrogen sink, storage of sulfur is controlled through the accumulation of specialized sulfur-rich proteins in maize endosperm. Silencing the synthesis of α-zeins through RNAi fails to raise sulfur-rich proteins. Although overexpression of the methionine-rich δ-zein can increase the methionine level in seeds, it occurs at least in part at the expense of the cysteine-rich β- and γ-zeins, demonstrating a balance between cysteine and methionine in sulfur storage. Therefore, we propose that the throttle for the flow of sulfur is placed before the synthesis of sulfur amino acids when sulfur is taken up and reduced during photosynthesis.
大多数开花植物种子由胚和胚乳组成,它们被母体组织包围,尤其是种皮。胚是处于休眠状态的后代,而胚乳则是分别用于储存蛋白质中的糖和碳水化合物中的氨基酸的终末器官。糖和氨基酸在光合作用期间于母体叶片中产生,开花后被运输到发育中的种子,在种子萌发期间,它们为幼苗早期生长提供养分。玉米胚乳通常含有约10%的蛋白质和70%的淀粉,且它们的组成比例相当稳定,因为这是通过由许多相互作用或相互拮抗的基因及途径网络构建的预设遗传程序严格调控的。然而,胚乳蛋白的营养价值较低,主要原因是α - 玉米醇溶蛋白基因家族的高表达,该家族编码不含赖氨酸的蛋白质。在不透明2(o2)突变体和α - 玉米醇溶蛋白RNA干扰(RNAi)转基因种子中,这些蛋白质水平的降低被非玉米醇溶蛋白的增加所补偿,导致氮库重新平衡,并使种子中总蛋白水平或多或少保持恒定。对于蛋白质含量为30%的培育玉米种子,也观察到了玉米醇溶蛋白和非玉米醇溶蛋白的相同重新平衡。与氮库不同,硫的储存是通过玉米胚乳中积累特殊的富含硫的蛋白质来控制的。通过RNAi使α - 玉米醇溶蛋白的合成沉默并不能提高富含硫的蛋白质水平。尽管富含蛋氨酸的δ - 玉米醇溶蛋白的过表达可以增加种子中的蛋氨酸水平,但这至少部分是以富含半胱氨酸的β - 和γ - 玉米醇溶蛋白为代价的,这表明在硫储存中半胱氨酸和蛋氨酸之间存在平衡。因此,我们提出,当硫在光合作用期间被吸收和还原时,硫流的调节点位于硫氨基酸合成之前。