Jankowska-Steifer Ewa, Madej Maria, Niderla-Bielińska Justyna, Ruminski Sławomir, Flaht-Zabost Aleksandra, Czarnowska Elzbieta, Gula Grzegorz, Radomska-Leśniewska Dorota M, Ratajska Anna
Department of Histology and Embryology, Medical University of Warsaw, Warsaw, Poland,
Histochem Cell Biol. 2015 Feb;143(2):153-69. doi: 10.1007/s00418-014-1269-z. Epub 2014 Sep 9.
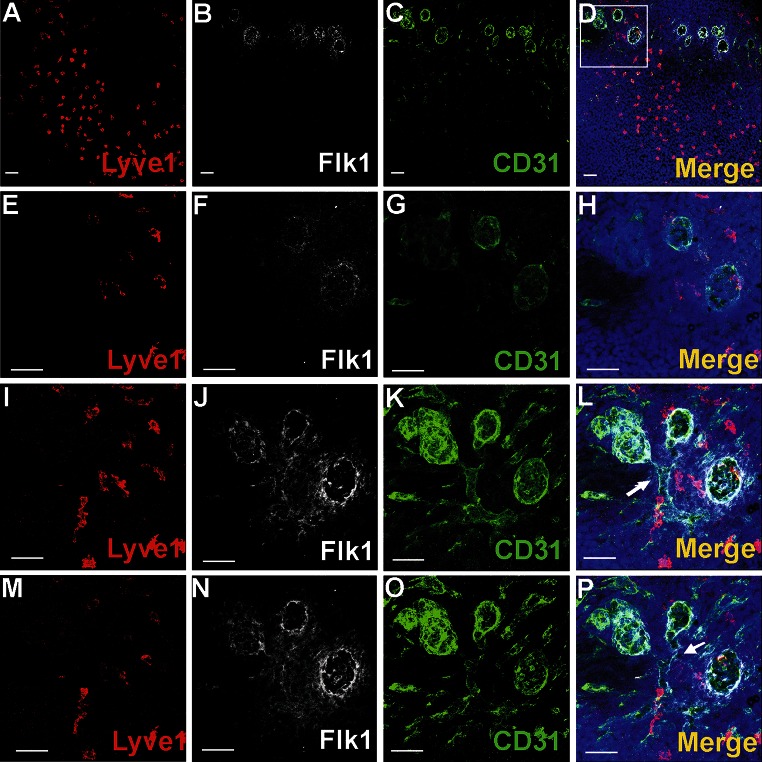
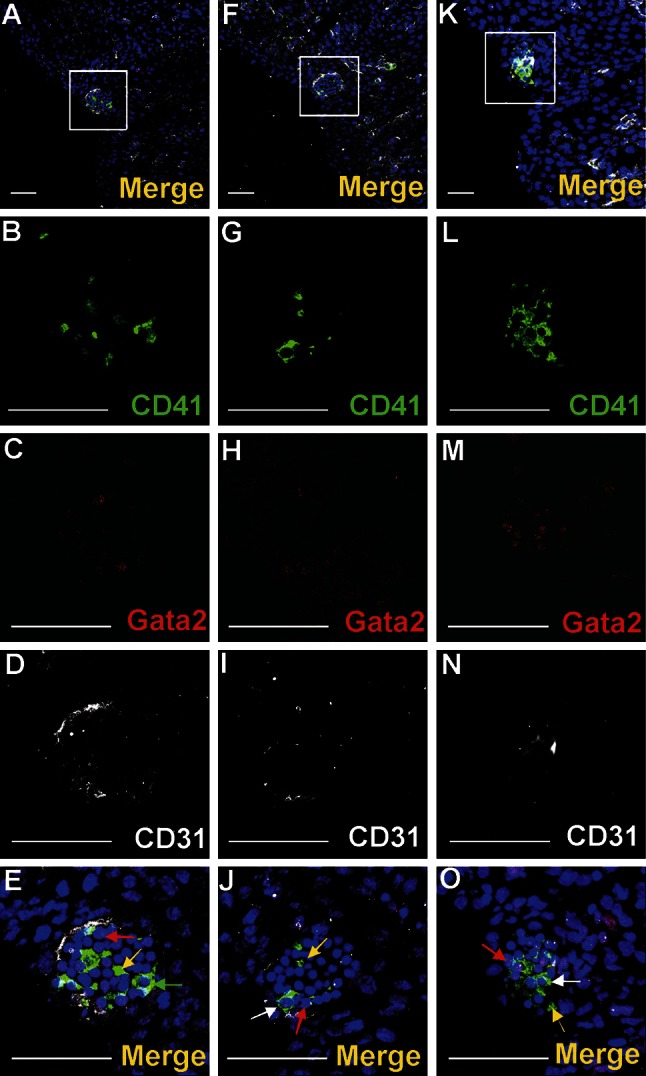
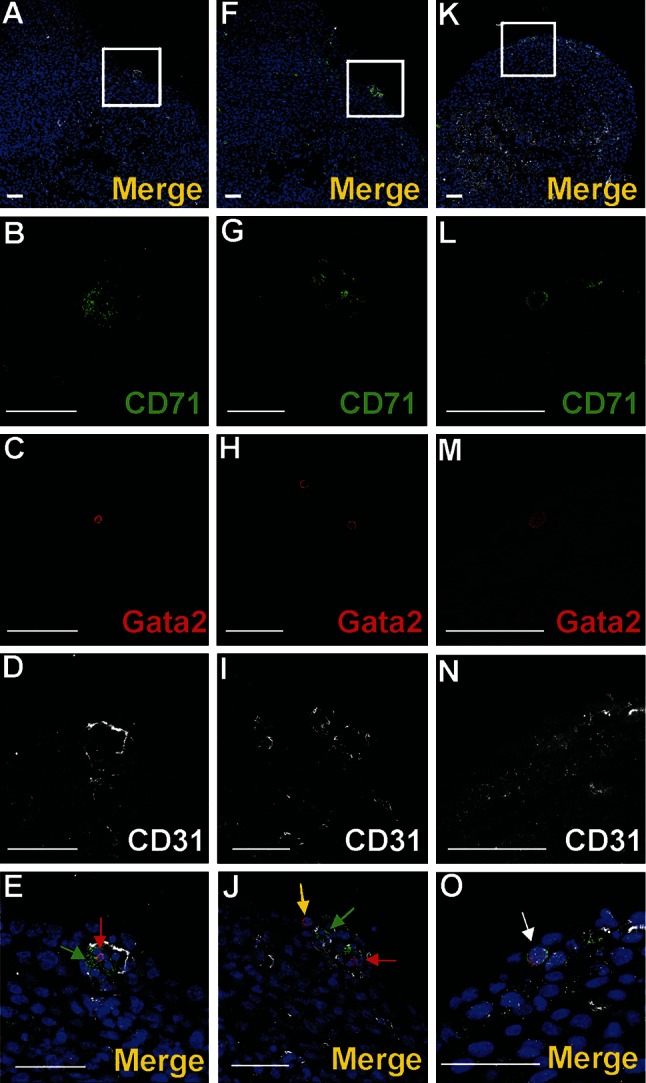
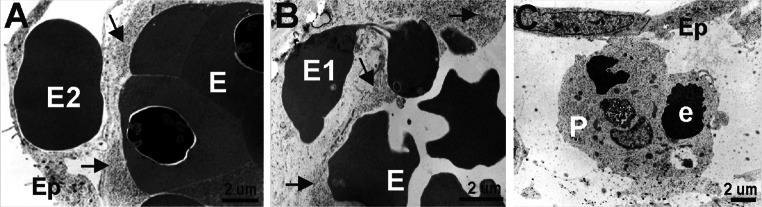
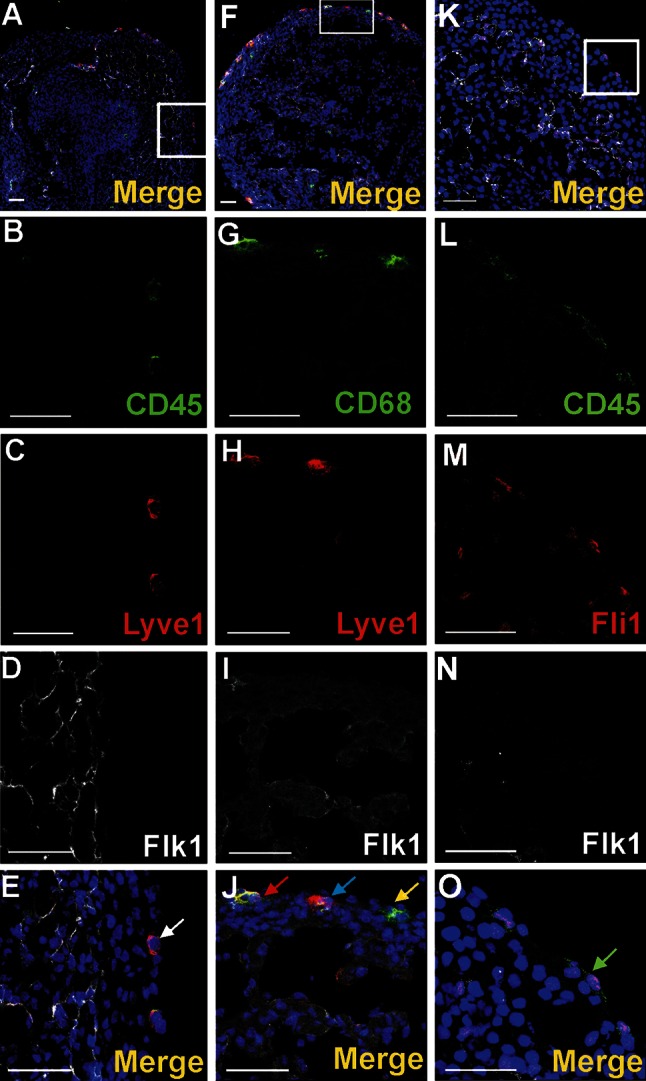
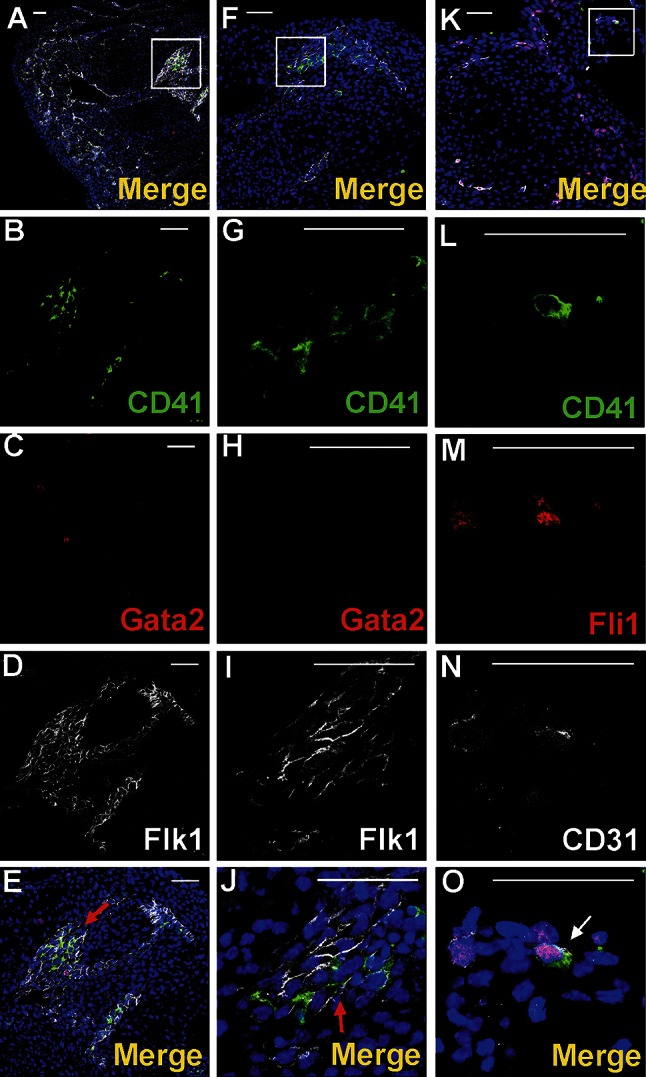
Vasculogenesis and hematopoiesis are co-localized in the embryonic body, but precise phenotypes of the cells contributing to these processes are not defined. The aim of this study was to characterize phenotypic profiles and location of putative vasculogenic and hematopoietic cellular progenitors in the embryonic mouse heart. Confocal microscopy, as well as ultrastructural and stereomicroscopic analyses, was performed on immunohistochemical whole-mount-stained or sectioned hearts at stages 11.5-14 dpc. A FASC analysis was conducted to quantify putative vasculogenic and hematopoietic cells. We found subepicardial blood islands in the form of foci of accumulation of cells belonging to erythroblastic and megakaryocytic lineages at various stages of maturation, exhibiting phenotypes: GATA2(+)/CD41(+), GATA2(-)/CD41(+), GATA2(+)/CD71(-), GATA2(-)/CD71(+), Fli1(+)/CD71(+), Fli1(-)/CD71(+), with a majority of cells expressing the Ter119 antigen, but none of them expressing Flk1. The subepicardium and the outflow tract endothelium were recognized to be the areas where progenitor cells were scattered or adjoining the endothelial cells. These progenitor cells were characterized as possessing the following antigens: CD45(+)/Fli1(+), CD41(+)/Flk1(+), Flk1(+)/Fli1(+). A FACS analysis demonstrated that the CD41/Flk1 double-positive population of cells constituted 2.68% of total cell population isolated from 12.5 dpc hearts. Vessels and tubules were positive for CD31, Flk1, Fli1, Tie2, including blood islands endothelia. The endocardial wall endothelia were found to function as an anchoring apparatus for megakaryocytes releasing platelets into the cardiac cavities. Phenotypic characteristics of vasculogenic (Flk1(+)/Fli1(+)) and hematopoietic (GATA2(+)/CD71(+), CD41(+)/GATA2(+)) progenitors, as well as the putative hemogenic endothelium (Flk1(+)/CD41(+)) in embryonic mouse hearts, have been presented. Cardiac blood islands, the subepicardium and endothelium of the outflow tract cushions have been defined as areas where these progenitor cells can be found.
血管生成和造血作用在胚胎体内共定位,但参与这些过程的细胞的精确表型尚未明确。本研究的目的是表征胚胎小鼠心脏中假定的血管生成和造血细胞祖细胞的表型特征及位置。对胚胎发育第11.5 - 14天的免疫组织化学全层染色或切片心脏进行共聚焦显微镜检查以及超微结构和立体显微镜分析。进行FASC分析以量化假定的血管生成和造血细胞。我们发现心外膜下血岛,其形式为处于不同成熟阶段的属于成红细胞和巨核细胞谱系的细胞聚集灶,表现出以下表型:GATA2(+)/CD41(+)、GATA2(-)/CD41(+)、GATA2(+)/CD71(-)、GATA2(-)/CD71(+)、Fli1(+)/CD71(+)、Fli1(-)/CD71(+),大多数细胞表达Ter119抗原,但均不表达Flk1。心外膜下层和流出道内皮被认为是祖细胞散布或与内皮细胞相邻的区域。这些祖细胞的特征是具有以下抗原:CD45(+)/Fli1(+)、CD41(+)/Flk1(+)、Flk1(+)/Fli1(+)。FACS分析表明,CD41/Flk1双阳性细胞群体占从胚胎发育第12.5天心脏分离的总细胞群体的2.68%。血管和小管对CD31、Flk1、Fli1、Tie2呈阳性,包括血岛内的内皮。发现心内膜壁内皮作为巨核细胞向心腔释放血小板的锚定装置。本文呈现了胚胎小鼠心脏中血管生成祖细胞(Flk1(+)/Fli1(+))和造血祖细胞(GATA2(+)/CD71(+)、CD41(+)/GATA2(+))以及假定的造血内皮(Flk1(+)/CD41(+))的表型特征。心脏血岛、心外膜下层和流出道垫的内皮已被定义为可发现这些祖细胞的区域。