Rodriguez Elizabeth, Nan Ruodan, Li Keying, Gor Jayesh, Perkins Stephen J
From the Department of Structural and Molecular Biology, Division of Biosciences, Darwin Building, University College London, Gower Street, London WC1E 6BT, United Kingdom.
From the Department of Structural and Molecular Biology, Division of Biosciences, Darwin Building, University College London, Gower Street, London WC1E 6BT, United Kingdom
J Biol Chem. 2015 Jan 23;290(4):2334-50. doi: 10.1074/jbc.M114.605691. Epub 2014 Dec 8.
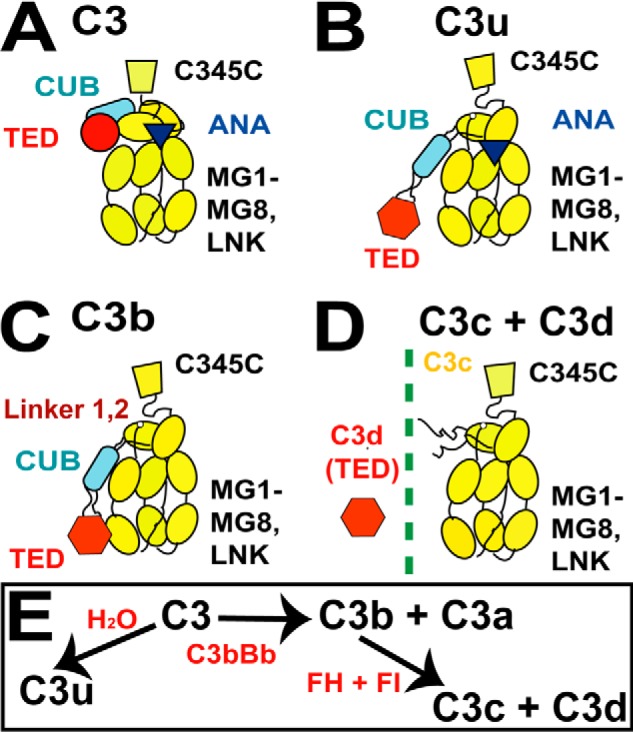
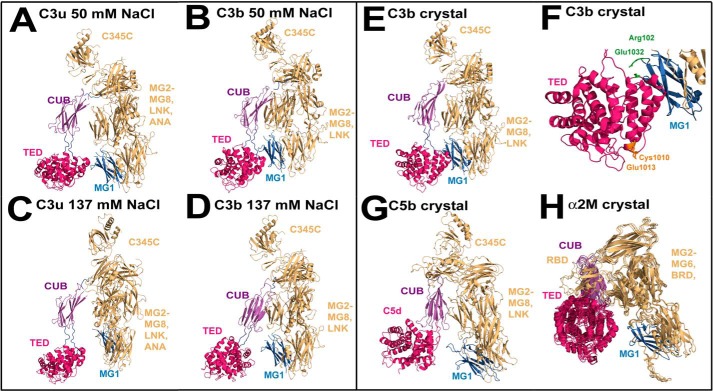
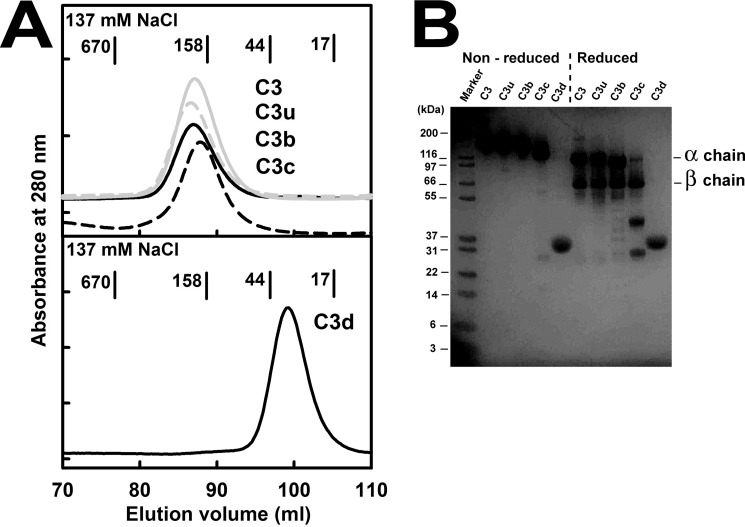
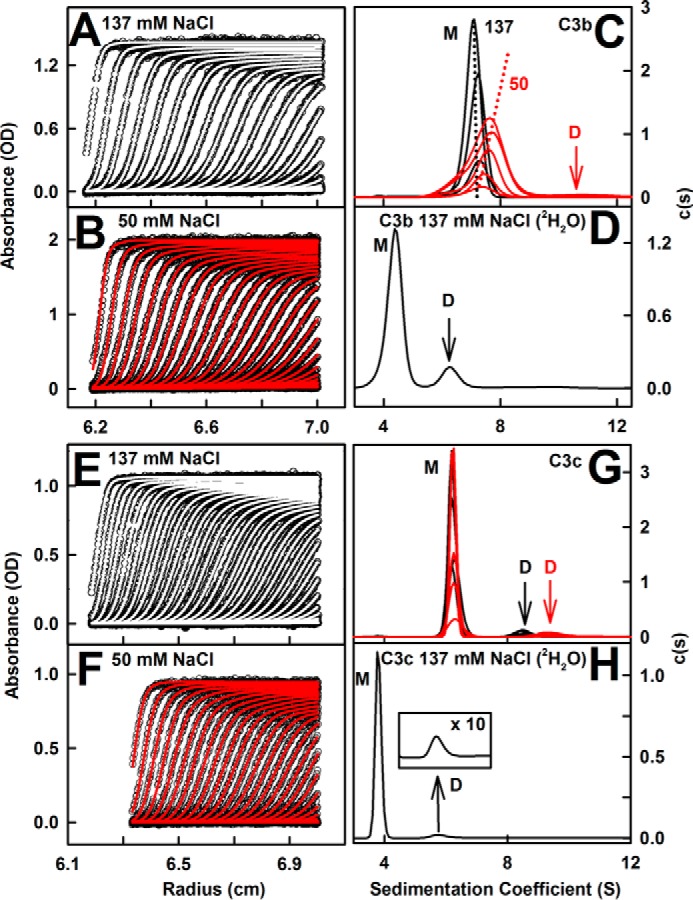
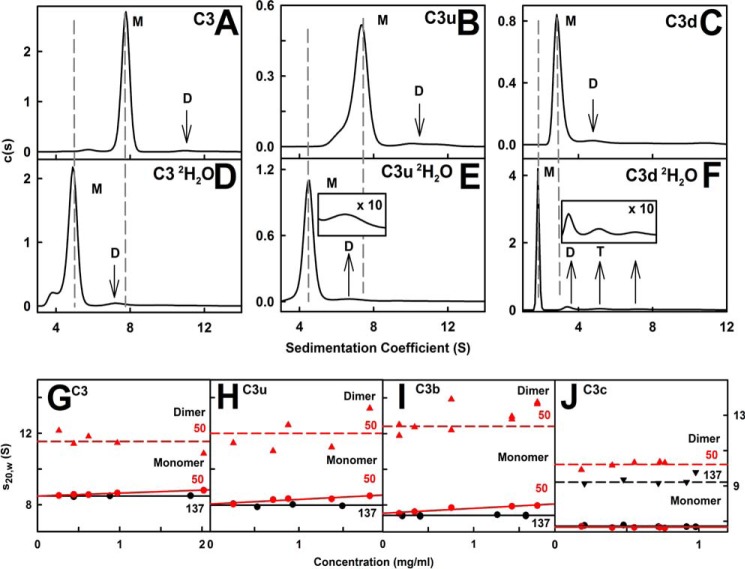
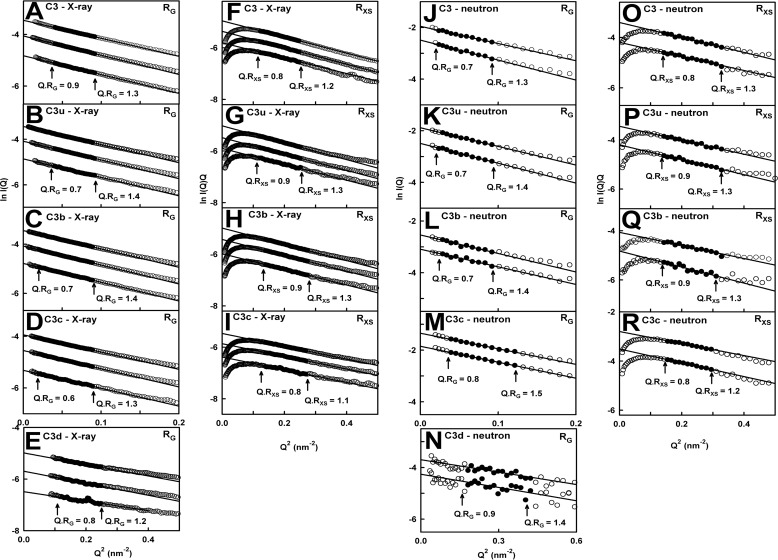
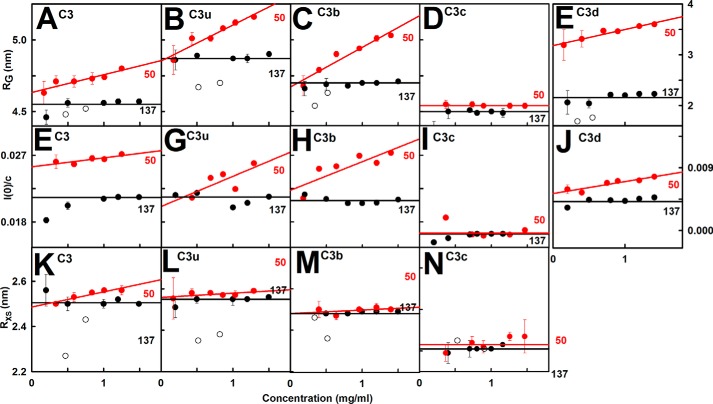
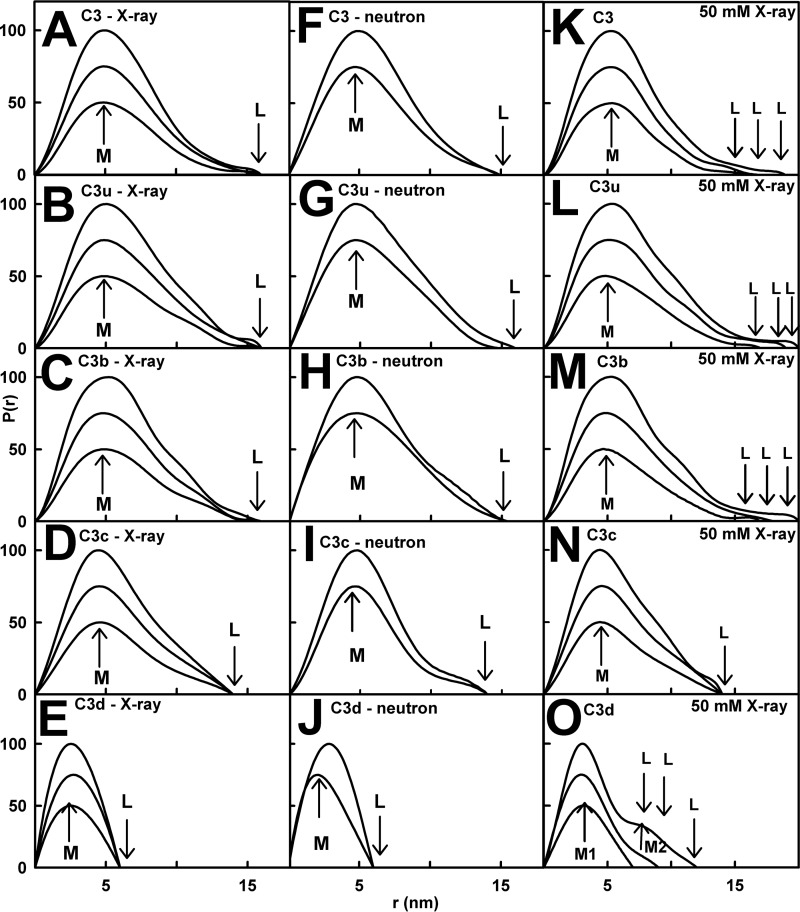
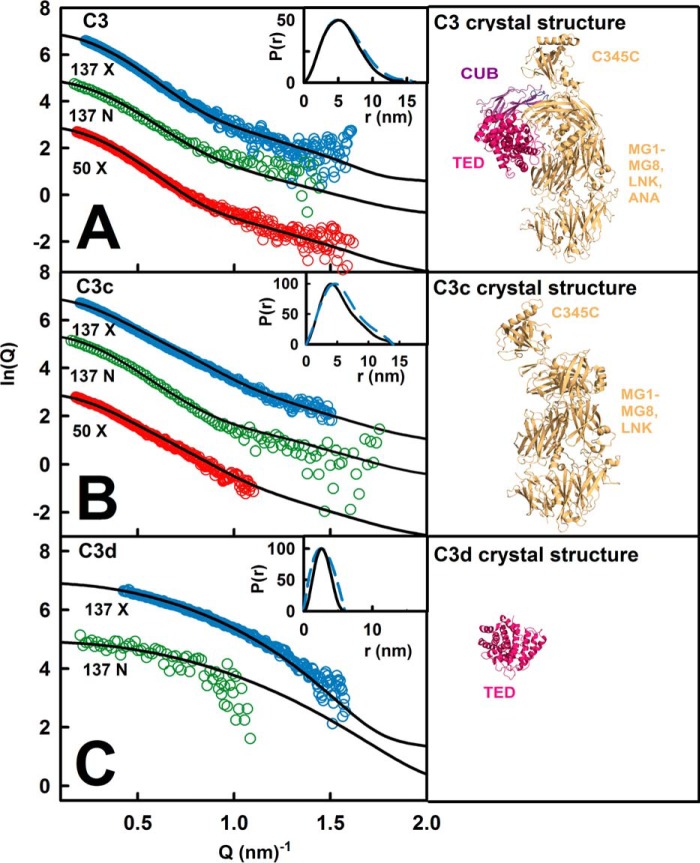
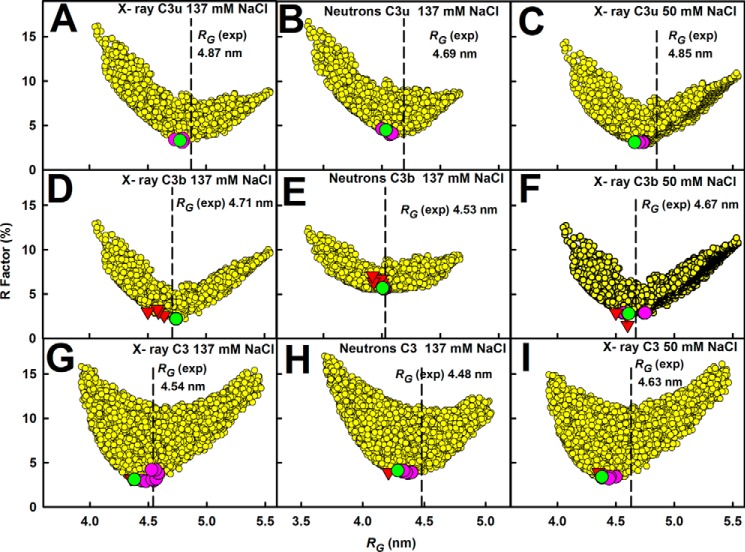
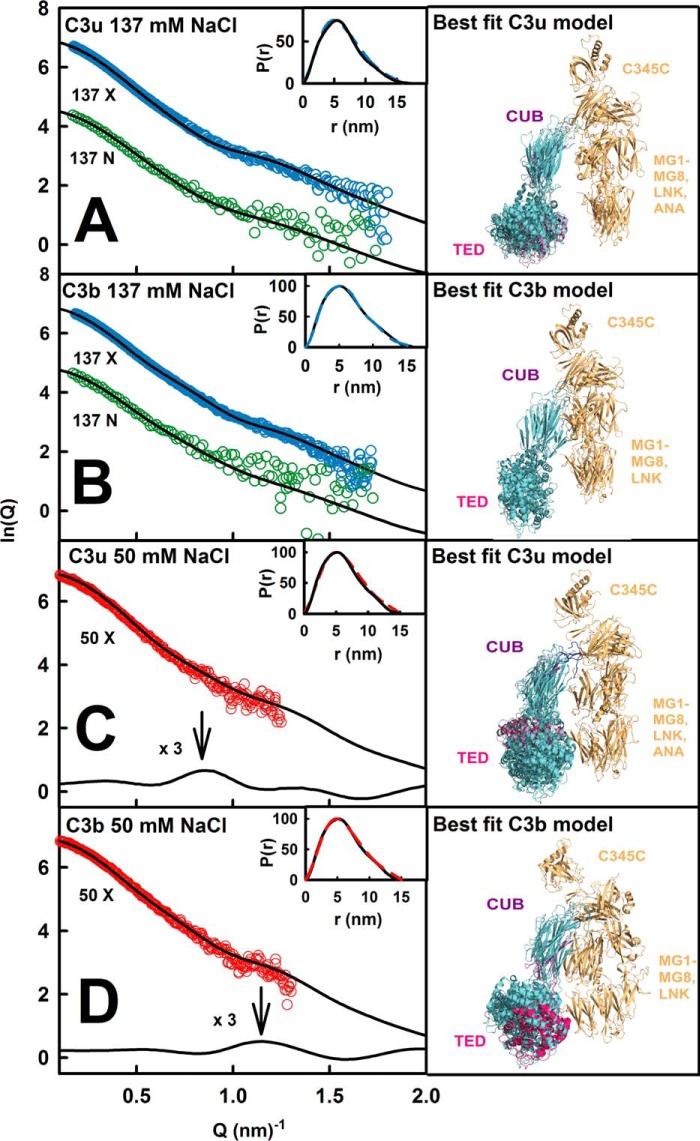
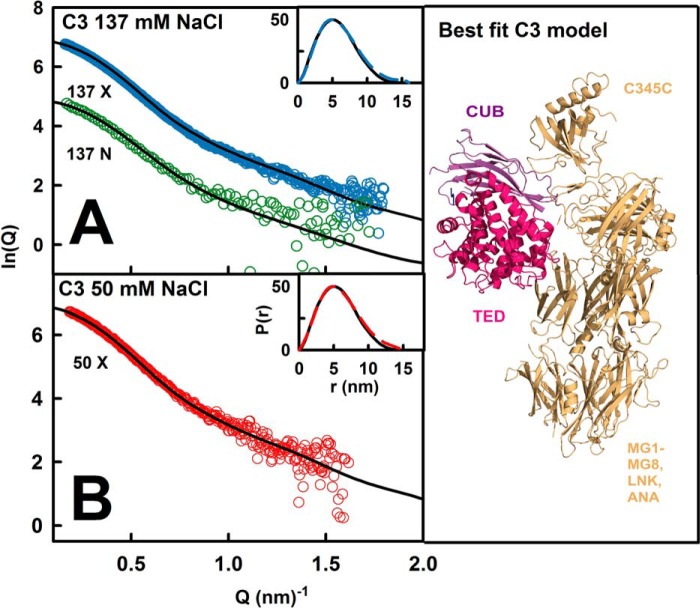
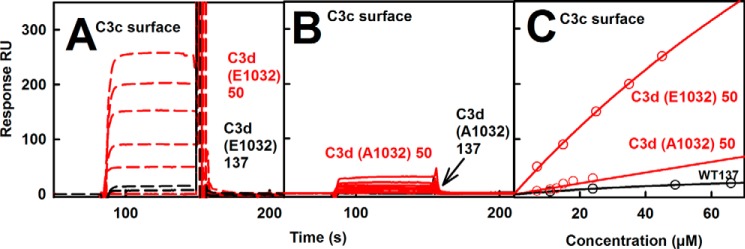
The solution structure of complement C3b is crucial for the understanding of complement activation and regulation. C3b is generated by the removal of C3a from C3. Hydrolysis of the C3 thioester produces C3u, an analog of C3b. C3b cleavage results in C3c and C3d (thioester-containing domain; TED). To resolve functional questions in relation to C3b and C3u, analytical ultracentrifugation and x-ray and neutron scattering studies were used with C3, C3b, C3u, C3c, and C3d, using the wild-type allotype with Arg(102). In 50 mm NaCl buffer, atomistic scattering modeling showed that both C3b and C3u adopted a compact structure, similar to the C3b crystal structure in which its TED and macroglobulin 1 (MG1) domains were connected through the Arg(102)-Glu(1032) salt bridge. In physiological 137 mm NaCl, scattering modeling showed that C3b and C3u were both extended in structure, with the TED and MG1 domains now separated by up to 6 nm. The importance of the Arg(102)-Glu(1032) salt bridge was determined using surface plasmon resonance to monitor the binding of wild-type C3d(E1032) and mutant C3d(A1032) to immobilized C3c. The mutant did not bind, whereas the wild-type form did. The high conformational variability of TED in C3b in physiological buffer showed that C3b is more reactive than previously thought. Because the Arg(102)-Glu(1032) salt bridge is essential for the C3b-Factor H complex during the regulatory control of C3b, the known clinical associations of the major C3S (Arg(102)) and disease-linked C3F (Gly(102)) allotypes of C3b were experimentally explained for the first time.
补体C3b的溶液结构对于理解补体激活和调节至关重要。C3b是通过从C3中去除C3a而产生的。C3硫酯键的水解产生C3u,它是C3b的类似物。C3b裂解产生C3c和C3d(含硫酯结构域;TED)。为了解决与C3b和C3u相关的功能问题,对野生型同种异型的C3、C3b、C3u、C3c和C3d进行了分析超速离心以及X射线和中子散射研究,该野生型同种异型含有精氨酸(102)。在50 mM NaCl缓冲液中,原子散射模型显示C3b和C3u均采用紧密结构,类似于C3b晶体结构,其中其TED和巨球蛋白1(MG1)结构域通过精氨酸(102)-谷氨酸(1032)盐桥相连。在生理浓度的137 mM NaCl中,散射模型显示C3b和C3u在结构上均呈伸展状态,此时TED和MG1结构域之间的距离可达6 nm。使用表面等离子体共振监测野生型C3d(E1032)和突变型C3d(A1032)与固定化C3c的结合,从而确定了精氨酸(102)-谷氨酸(1032)盐桥的重要性。突变体不结合,而野生型形式则结合。生理缓冲液中C3b的TED具有高度的构象变异性,这表明C3b比以前认为的更具反应性。由于在C3b的调节控制过程中,精氨酸(102)-谷氨酸(1032)盐桥对于C3b-因子H复合物至关重要,因此首次通过实验解释了C3b主要的C3S(精氨酸(102))和与疾病相关的C3F(甘氨酸(102))同种异型的已知临床关联。