Falck-Pedersen E, Logan J
Hearst Research Foundation, Department of Microbiology, Cornell University Medical College, New York, New York 10021.
J Virol. 1989 Feb;63(2):532-41. doi: 10.1128/JVI.63.2.532-541.1989.
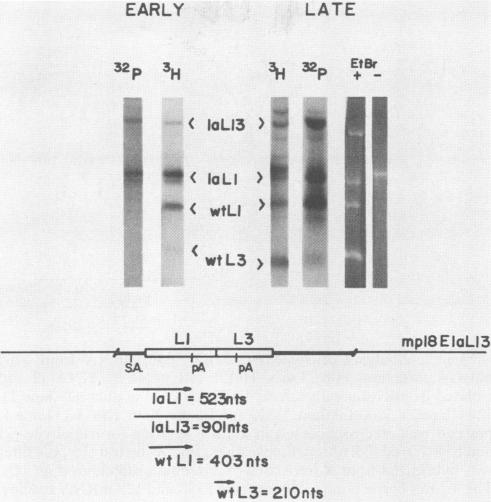
We have investigated the mechanisms involved in the early-to-late RNA-processing switch which regulates the mRNA species generated from the adenovirus major late transcription unit (MLTU). In particular, polyadenylation choice mechanisms were characterized by using a reconstructed adenovirus E1A gene as a site for insertion of MLTU poly(A) regulation signals (L1 and L3). Adenovirus constructs containing the variant poly(A) recognition elements were used to compare E1A poly(A) signal utilization with wild-type MLTU (L1 to L5) utilization. In both early and late stages of infection, either polyadenylation site (L1 or L3) is capable of being utilized when presented as the only operational poly(A) site. In an early infection, a virus which contains multiple elements presented in tandem (L13) uses the first poly(A) site, L1, preferentially (ratio of L1 to L3, 8:1) in both E1A and MLTU loci. Transcription termination is not involved in restricting the utilization of the downstream L3 site. In a late infection, when each of the five MLTU poly(A) sites is used, a switch also occurs for the E1AL13 construct, with utilization of both the L1 and L3 poly(A) sites. The switch from early to late was not the result of altered processing factors in the late infection, as demonstrated by superinfecting the E1AL13 construct into cells which had already entered a late stage of infection. The superinfecting virus gave an L1-only phenotype; therefore, a cis mechanism is involved in adenovirus poly(A) regulation.
我们研究了参与早期到晚期RNA加工转换的机制,该转换调节由腺病毒主要晚期转录单位(MLTU)产生的mRNA种类。具体而言,通过使用重建的腺病毒E1A基因作为插入MLTU聚腺苷酸化(poly(A))调控信号(L1和L3)的位点,对聚腺苷酸化选择机制进行了表征。含有变异聚(A)识别元件的腺病毒构建体用于比较E1A聚(A)信号利用与野生型MLTU(L1至L5)利用情况。在感染的早期和晚期阶段,当作为唯一可操作的聚(A)位点呈现时,任一聚腺苷酸化位点(L1或L3)都能够被利用。在早期感染中,一种串联呈现多个元件(L13)的病毒在E1A和MLTU基因座中优先使用第一个聚(A)位点L1(L1与L3的比例为8:1)。转录终止不参与限制下游L3位点的利用。在晚期感染中,当使用五个MLTU聚(A)位点中的每一个时,E1AL13构建体也会发生转换,同时利用L1和L3聚(A)位点。从早期到晚期的转换不是晚期感染中加工因子改变的结果,这通过将E1AL13构建体超感染到已经进入感染晚期的细胞中得到证明。超感染的病毒呈现仅L1的表型;因此,腺病毒聚(A)调控涉及一种顺式机制。