O'Brien Martin, Kaplan-Levy Ruth N, Quon Tezz, Sappl Pia G, Smyth David R
School of Biological Sciences, Monash University, Melbourne, Vic. 3800, Australia.
School of Biological Sciences, Monash University, Melbourne, Vic. 3800, Australia
J Exp Bot. 2015 May;66(9):2475-85. doi: 10.1093/jxb/erv032. Epub 2015 Feb 19.
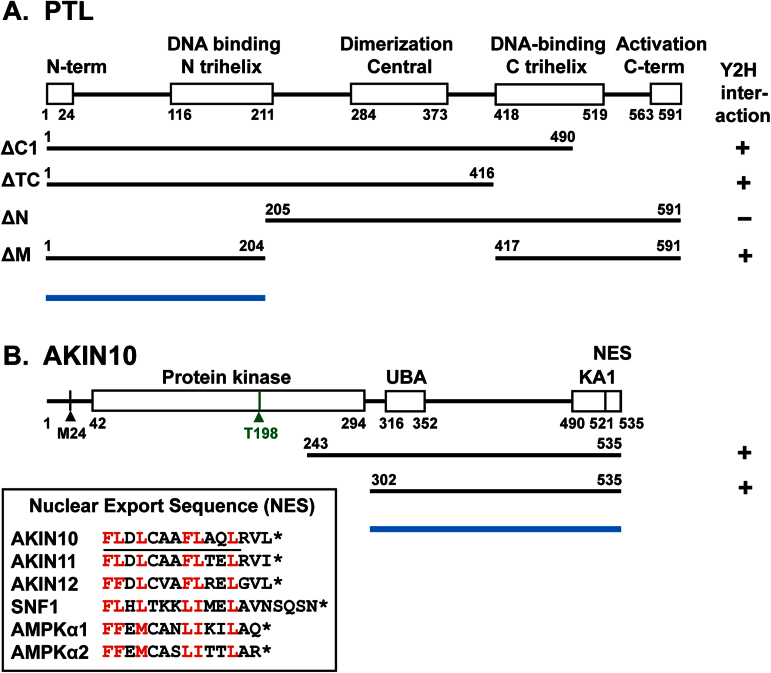
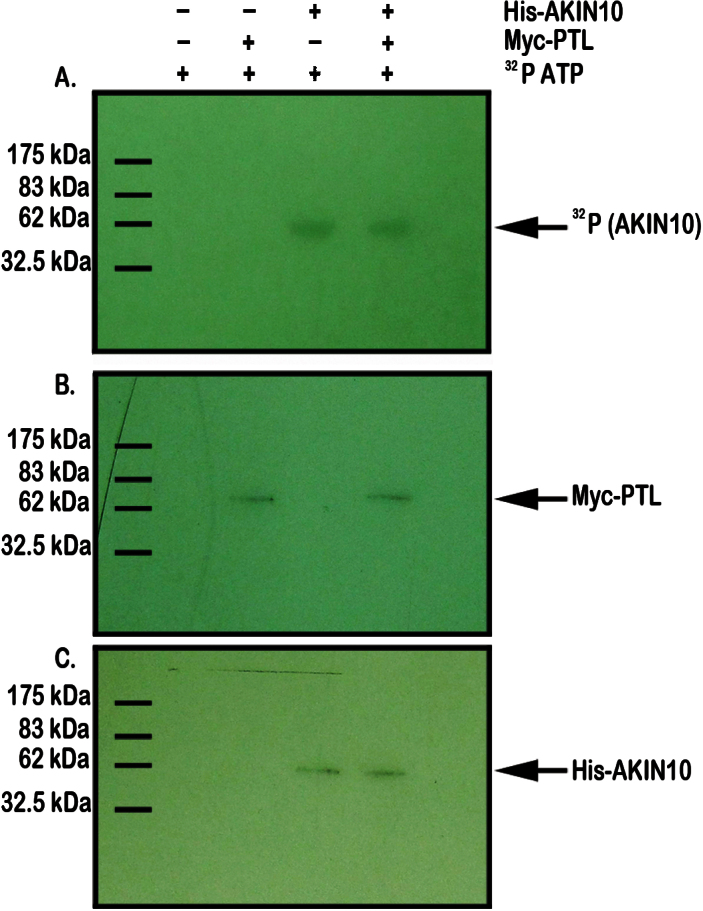
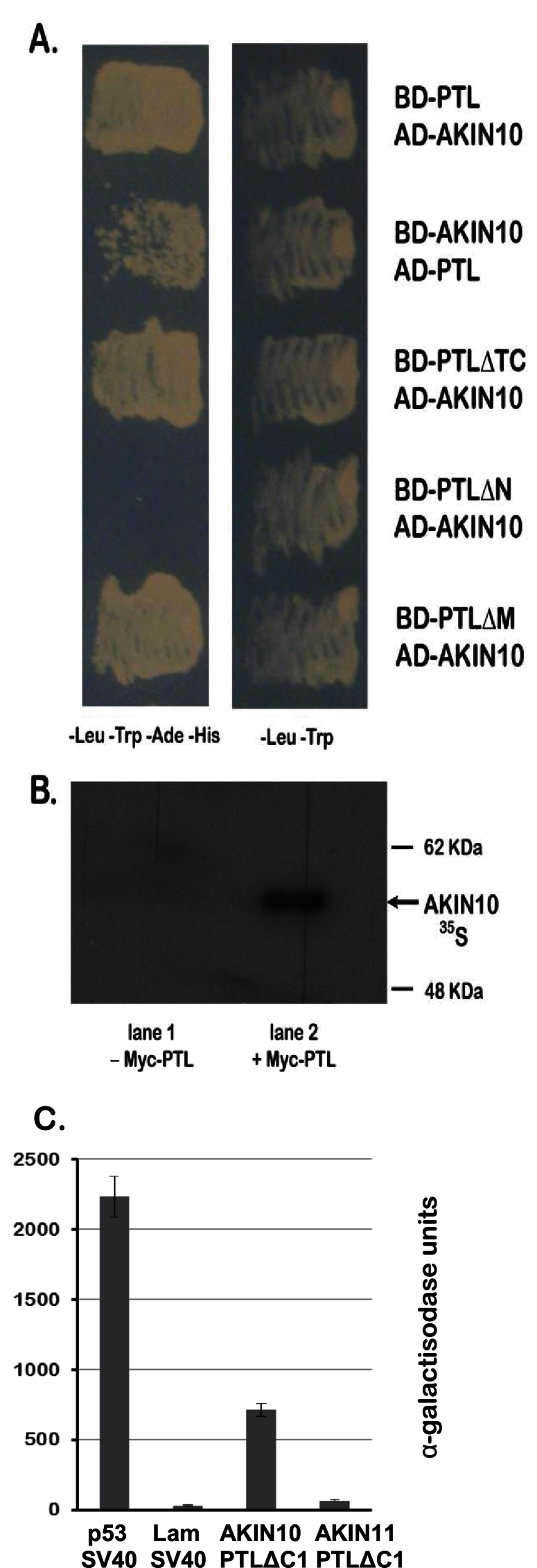
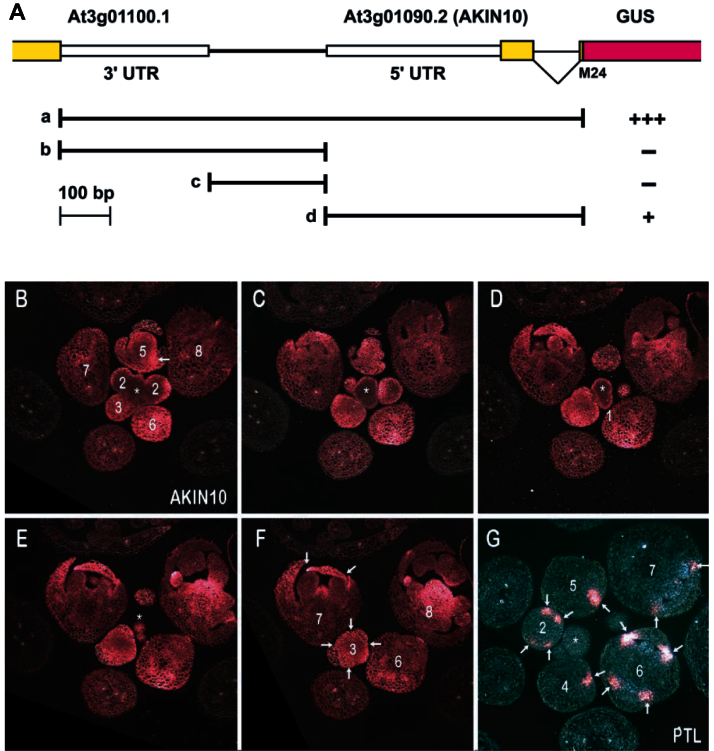
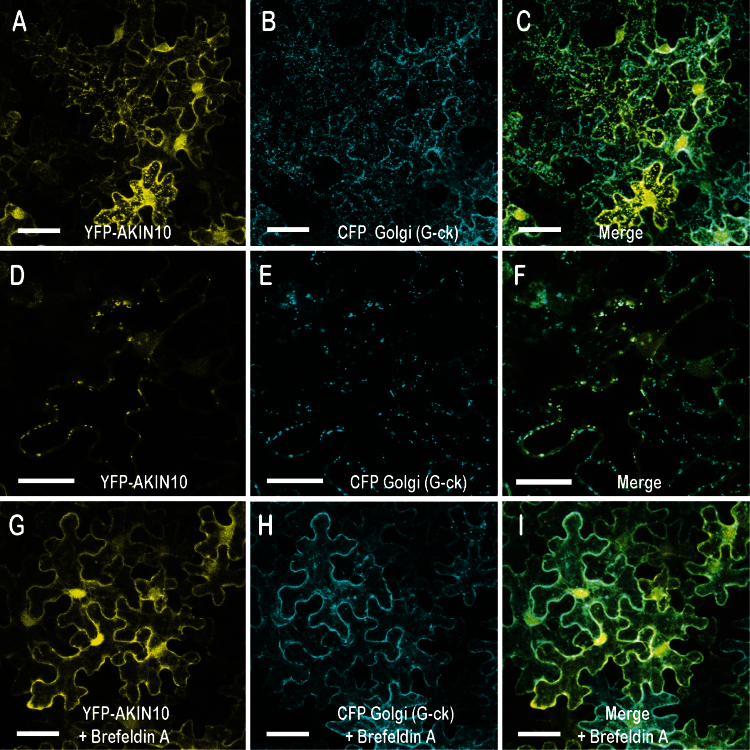
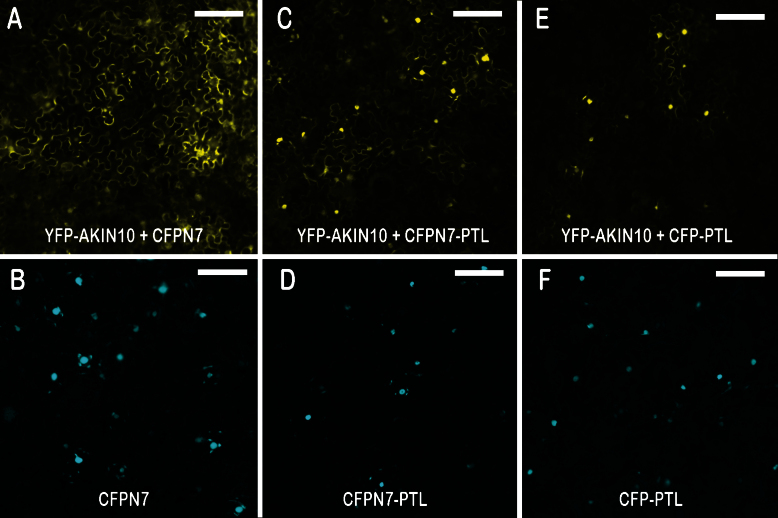
Organogenesis in plants involves differential growth. Rapidly growing primordia are distinguished from the meristem and each other by slower growing boundaries. PETAL LOSS (PTL) is a trihelix transcription factor of Arabidopsis that represses growth in boundaries between newly arising sepals. To identify partners involved in this growth limitation, a young inflorescence cDNA library was screened by yeast two-hybrid technology with PTL as bait. The most frequent prey identified was AKIN10, the catalytic α-subunit of the Snf1-related kinase1 (SnRK1). Interaction was mapped to the C-terminal (non-kinase) half of AKIN10 and the N-terminal portion of PTL. Binding of PTL was specific to AKIN10 as there was little binding to the related AKIN11. The interaction was confirmed by co-immunoprecipitation in vitro. Fluorescently tagged products of 35S:YFP-AKIN10 and 35S:CFP-PTL also interacted when transiently expressed together in leaf cells of Nicotiana benthamiana. In this case, most of the cytoplasmic AKIN10 was preferentially moved to the nucleus where PTL accumulated, possibly because a nuclear export sequence in AKIN10 was now masked. During these experiments, we observed that AKIN10 could variably accumulate in the Golgi, shown by its co-localization with a tagged Golgi marker and through its dispersal by brefeldin A. Tests of phosphorylation of PTL by AKIN10 gave negative results. The functional significance of the PTL-AKIN10 interaction remains open, although a testable hypothesis is that AKIN10 senses lower energy levels in inter-sepal zones and, in association with PTL, promotes reduced cell division.
植物器官发生涉及差异生长。快速生长的原基通过生长较慢的边界与分生组织以及彼此区分开来。花瓣缺失(PTL)是拟南芥中的一种三螺旋转录因子,它抑制新出现的萼片之间边界处的生长。为了鉴定参与这种生长限制的伙伴,以PTL为诱饵,通过酵母双杂交技术筛选了一个幼嫩花序cDNA文库。鉴定出的最常见猎物是AKIN10,即Snf1相关激酶1(SnRK1)的催化α亚基。相互作用定位于AKIN10的C末端(非激酶)一半和PTL的N末端部分。PTL与AKIN10的结合具有特异性,因为与相关的AKIN11几乎没有结合。通过体外共免疫沉淀证实了这种相互作用。当在本氏烟草的叶细胞中瞬时共表达时,35S:YFP - AKIN10和35S:CFP - PTL的荧光标记产物也相互作用。在这种情况下,大部分细胞质中的AKIN10优先转移到PTL积累的细胞核中,这可能是因为AKIN10中的核输出序列现在被掩盖了。在这些实验中,我们观察到AKIN10可以在高尔基体中不同程度地积累,这通过它与标记的高尔基体标志物的共定位以及通过布雷菲德菌素A使其分散来表明。AKIN10对PTL磷酸化的测试结果为阴性。尽管一个可检验的假设是AKIN10感知萼片间区域较低的能量水平,并与PTL一起促进细胞分裂减少,但PTL - AKIN10相互作用的功能意义仍不明确。