Vaisman Alexandra, Woodgate Roger
Laboratory of Genomic Integrity, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD 20892-3371, USA.
Laboratory of Genomic Integrity, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD 20892-3371, USA.
DNA Repair (Amst). 2015 May;29:74-82. doi: 10.1016/j.dnarep.2015.02.008. Epub 2015 Feb 16.
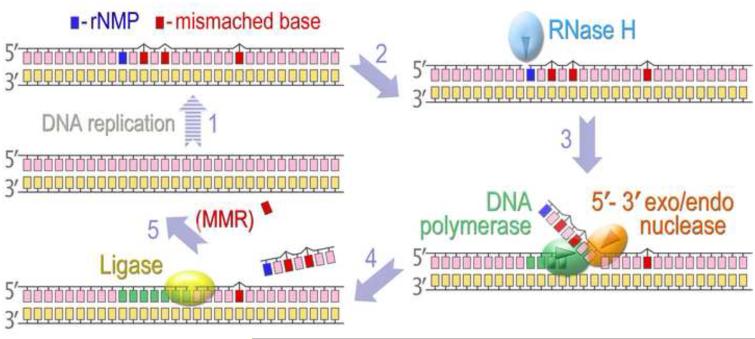
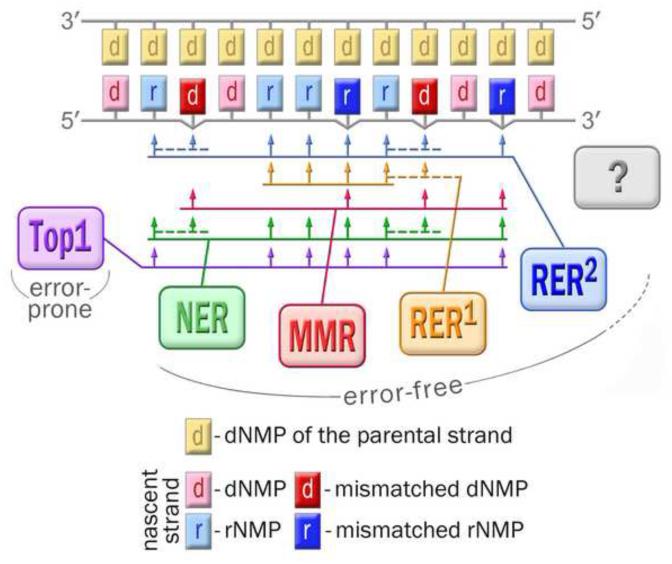
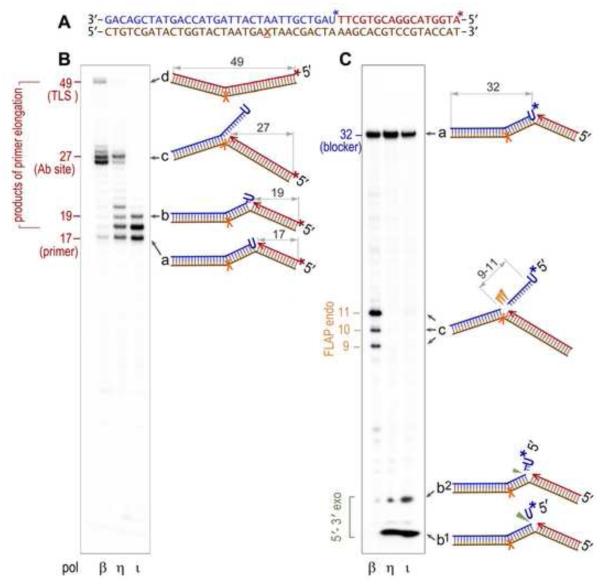
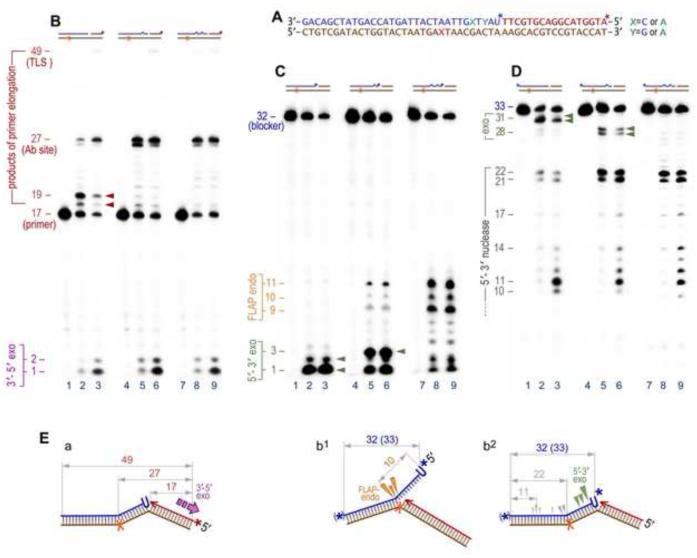
The survival of all living organisms is determined by their ability to reproduce, which in turn depends on accurate duplication of chromosomal DNA. In order to ensure the integrity of genome duplication, DNA polymerases are equipped with stringent mechanisms by which they select and insert correctly paired nucleotides with a deoxyribose sugar ring. However, this process is never 100% accurate. To fix occasional mistakes, cells have evolved highly sophisticated and often redundant mechanisms. A good example is mismatch repair (MMR), which corrects the majority of mispaired bases and which has been extensively studied for many years. On the contrary, pathways leading to the replacement of nucleotides with an incorrect sugar that is embedded in chromosomal DNA have only recently attracted significant attention. This review describes progress made during the last few years in understanding such pathways in both prokaryotes and eukaryotes. Genetic studies in Escherichia coli and Saccharomyces cerevisiae demonstrated that MMR has the capacity to replace errant ribonucleotides, but only when the base is mispaired. In contrast, the major evolutionarily conserved ribonucleotide repair pathway initiated by the ribonuclease activity of type 2 Rnase H has broad specificity. In yeast, this pathway also requires the concerted action of Fen1 and pol δ, while in bacteria it can be successfully completed by DNA polymerase I. Besides these main players, all organisms contain alternative enzymes able to accomplish the same tasks, although with differing efficiency and fidelity. Studies in bacteria have very recently demonstrated that isolated rNMPs can be removed from genomic DNA by error-free nucleotide excision repair (NER), while studies in yeast suggest the involvement of topoisomerase 1 in alternative mutagenic ribonucleotide processing. This review summarizes the most recent progress in understanding the ribonucleotide repair mechanisms in prokaryotes and eukaryotes.
所有生物的生存都取决于它们的繁殖能力,而繁殖能力又依赖于染色体DNA的精确复制。为确保基因组复制的完整性,DNA聚合酶具备严格的机制,通过这些机制它们选择并插入带有脱氧核糖糖环的正确配对核苷酸。然而,这一过程并非100%准确。为修正偶尔出现的错误,细胞进化出了高度复杂且往往冗余的机制。一个很好的例子是错配修复(MMR),它能纠正大多数错配碱基,并且已经被广泛研究多年。相反,导致嵌入染色体DNA中带有错误糖的核苷酸被替换的途径直到最近才引起广泛关注。这篇综述描述了过去几年在理解原核生物和真核生物中此类途径方面取得的进展。大肠杆菌和酿酒酵母中的遗传学研究表明,MMR有能力替换错误的核糖核苷酸,但仅当碱基错配时。相比之下,由2型核糖核酸酶H的核糖核酸酶活性引发的主要进化保守核糖核苷酸修复途径具有广泛的特异性。在酵母中,这条途径还需要Fen1和聚合酶δ的协同作用,而在细菌中它可以由DNA聚合酶I成功完成。除了这些主要参与者外,所有生物都含有能够完成相同任务的替代酶,尽管效率和保真度有所不同。细菌研究最近表明,游离的核糖核苷酸单磷酸(rNMPs)可以通过无差错核苷酸切除修复(NER)从基因组DNA中去除,而酵母研究表明拓扑异构酶1参与了替代性诱变核糖核苷酸加工。这篇综述总结了在理解原核生物和真核生物核糖核苷酸修复机制方面的最新进展。