Narasimhan Kamesh, Lambert Samuel A, Yang Ally W H, Riddell Jeremy, Mnaimneh Sanie, Zheng Hong, Albu Mihai, Najafabadi Hamed S, Reece-Hoyes John S, Fuxman Bass Juan I, Walhout Albertha J M, Weirauch Matthew T, Hughes Timothy R
Donnelly Centre for Cellular and Biomolecular Research, University of Toronto, Toronto, Canada.
Department of Molecular Genetics, University of Toronto, Toronto, Canada.
Elife. 2015 Apr 23;4:e06967. doi: 10.7554/eLife.06967.
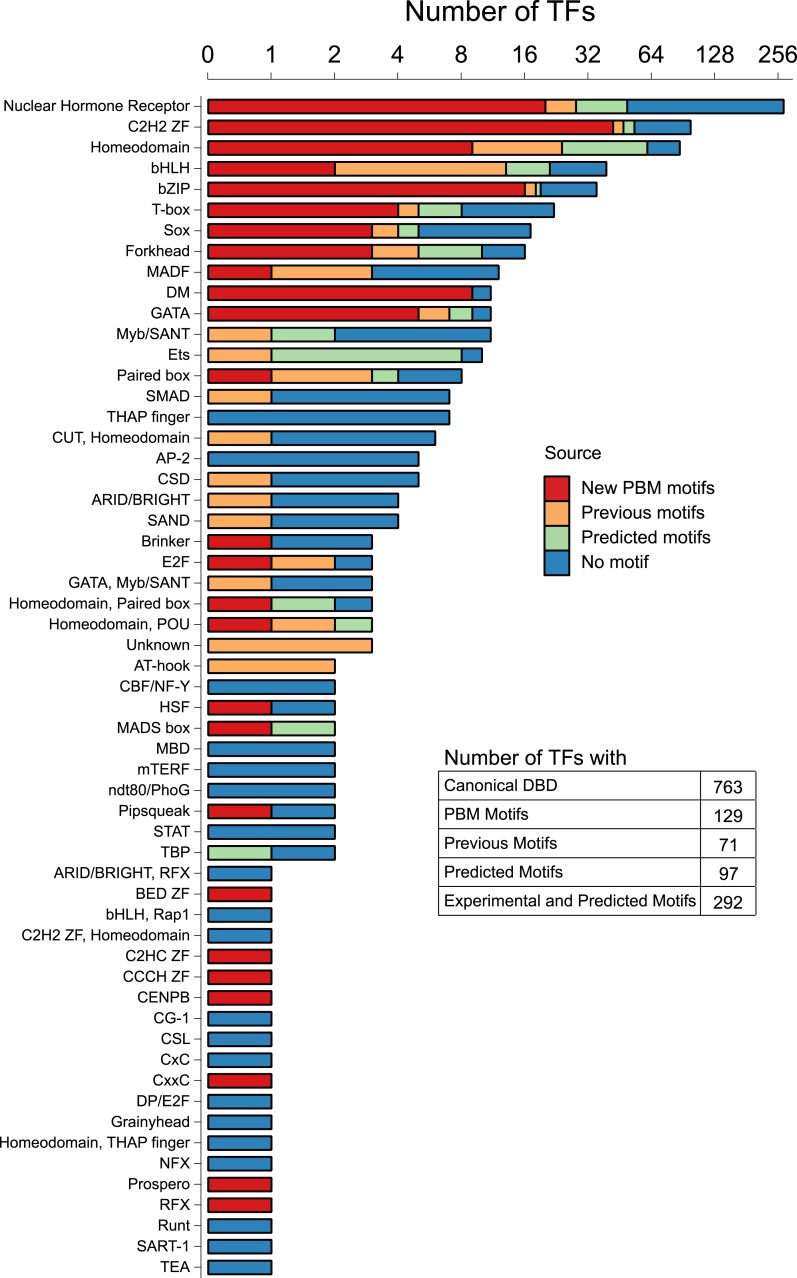
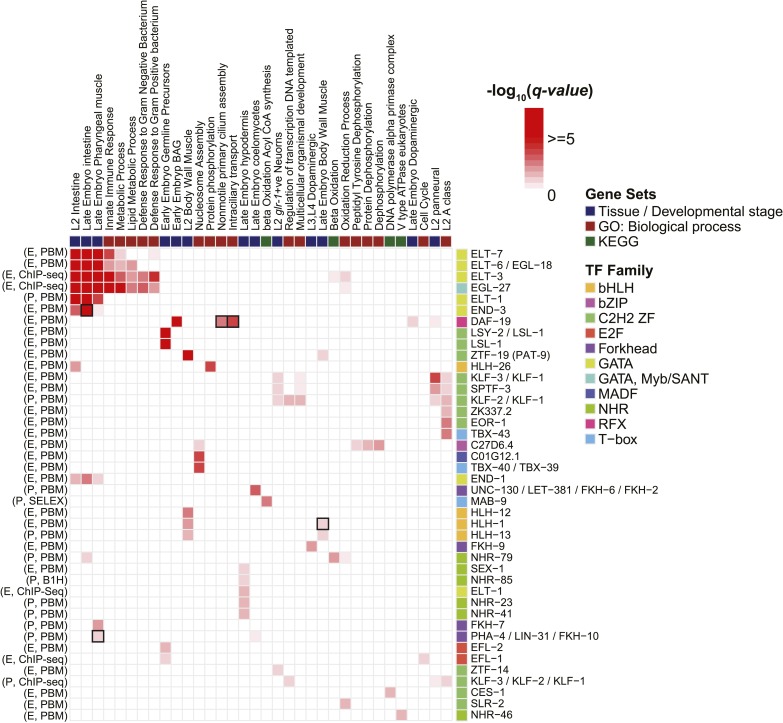
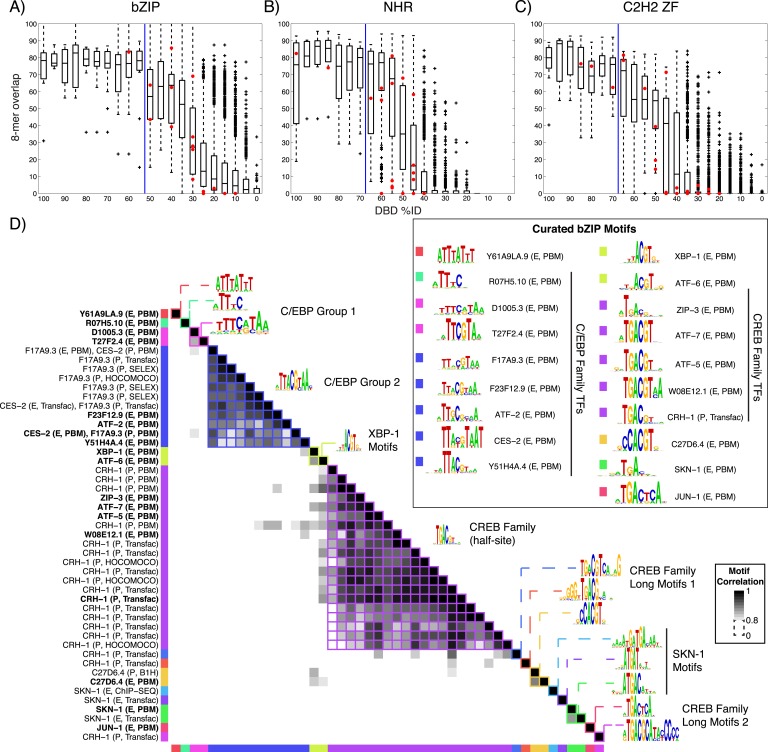
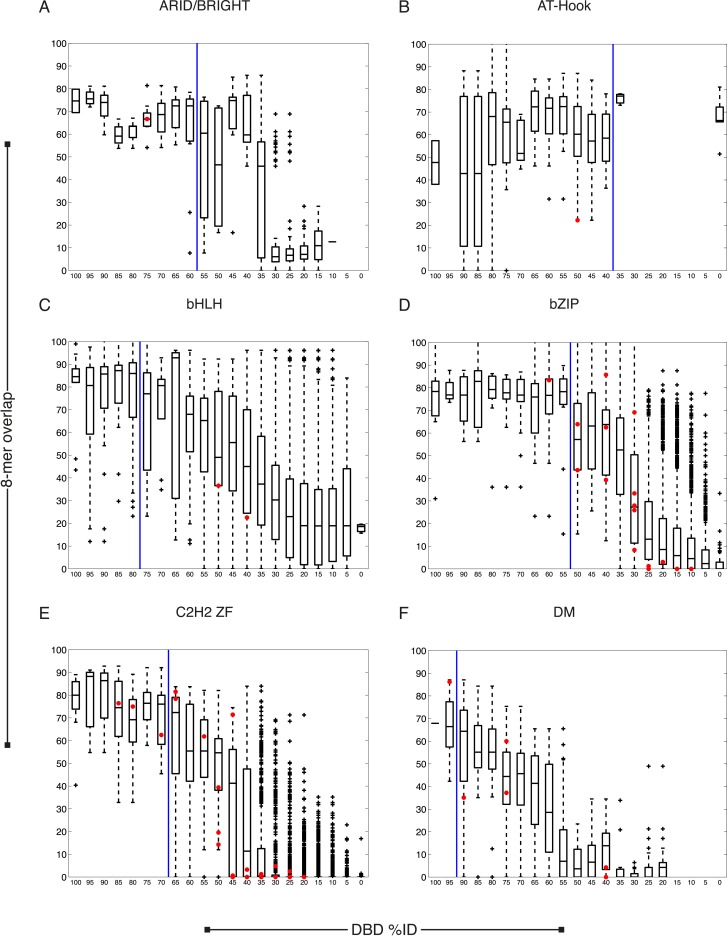
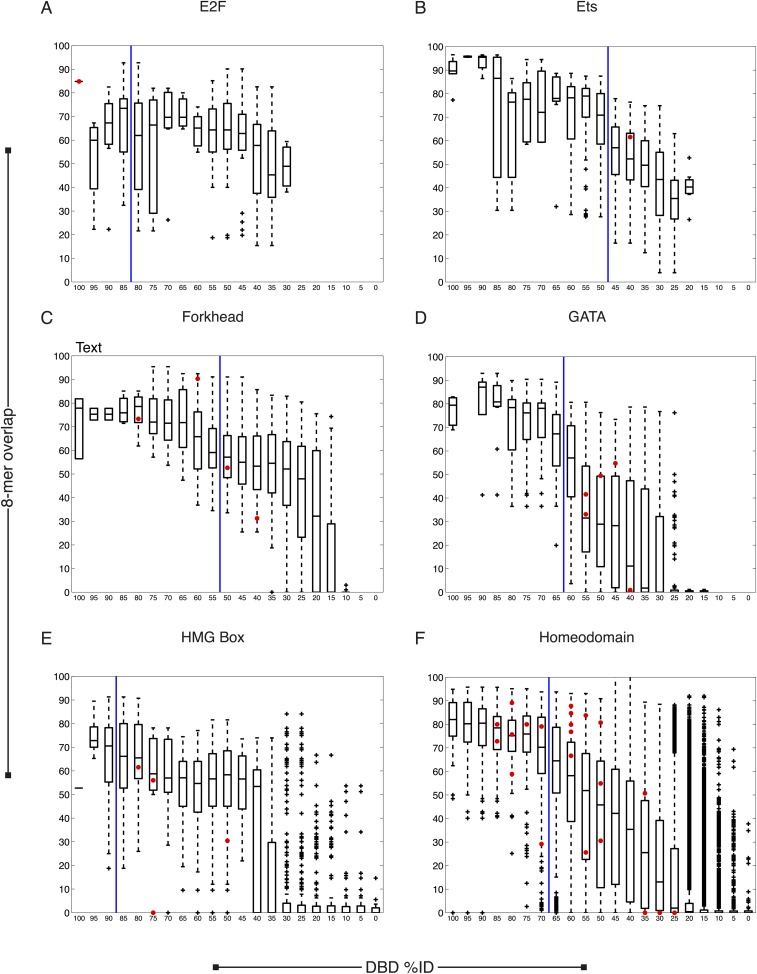
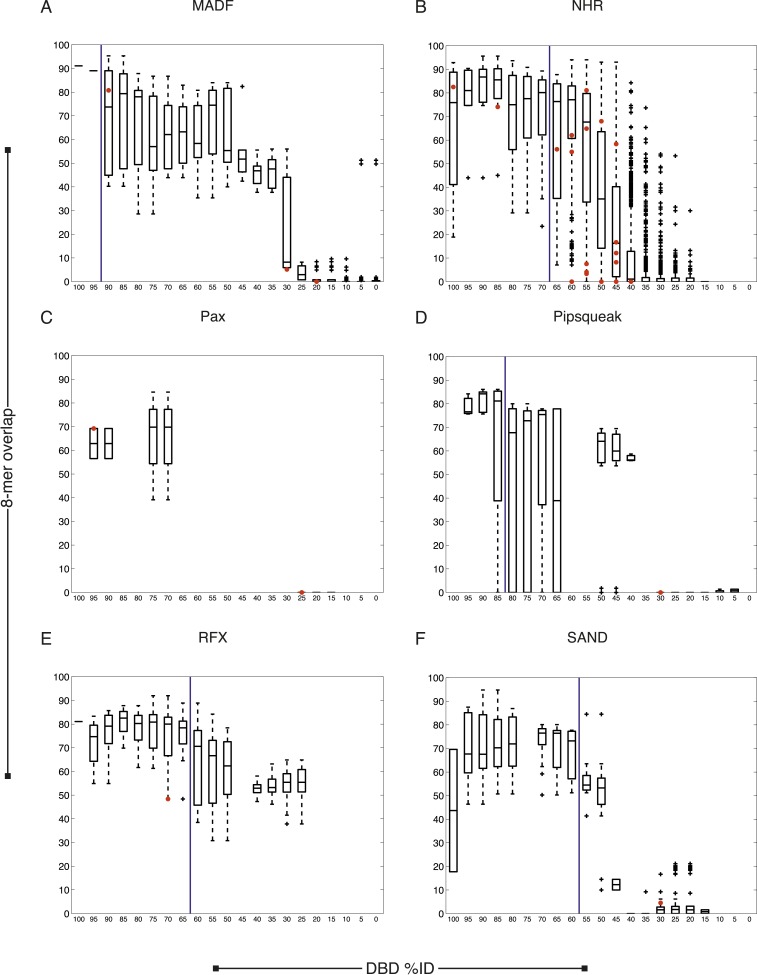
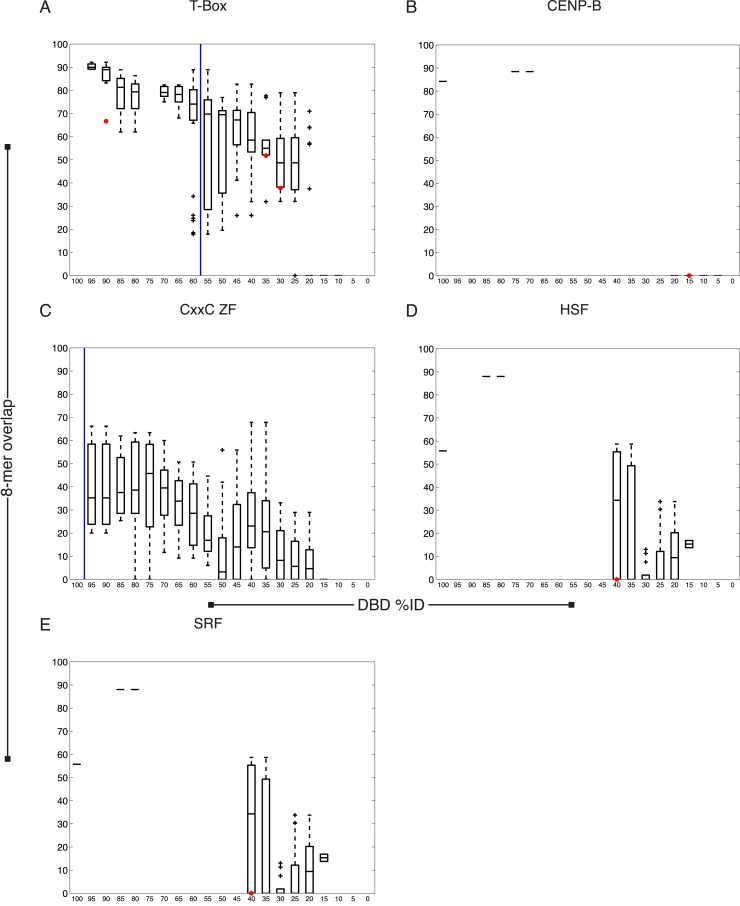
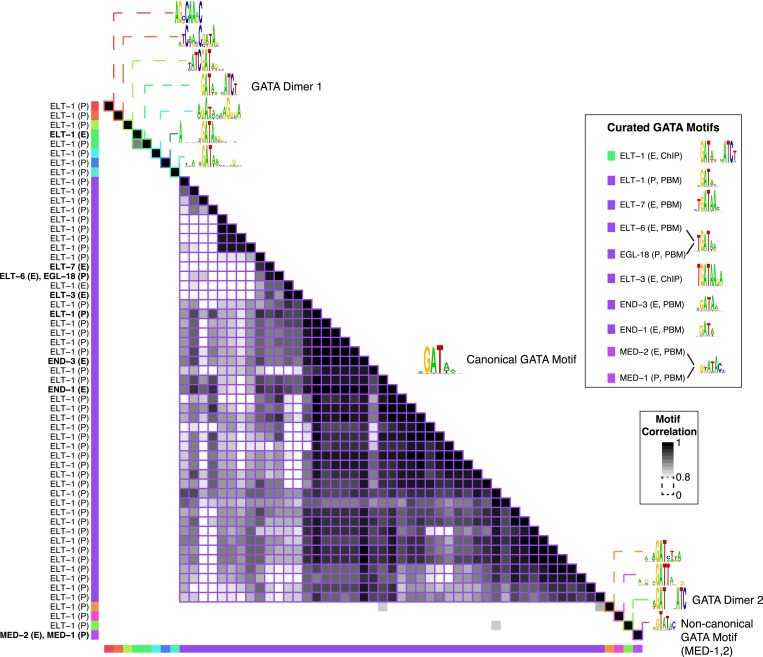
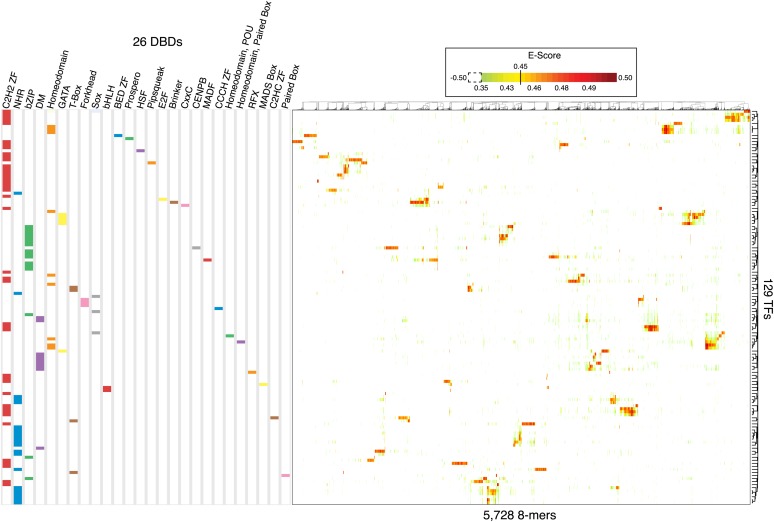
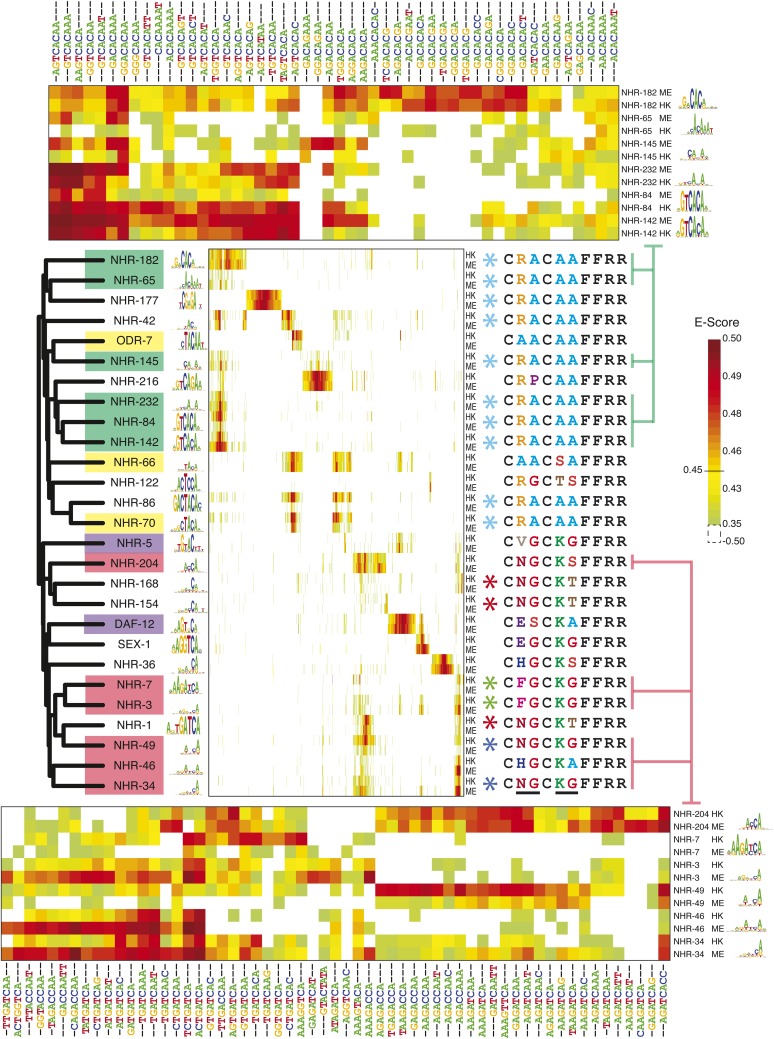
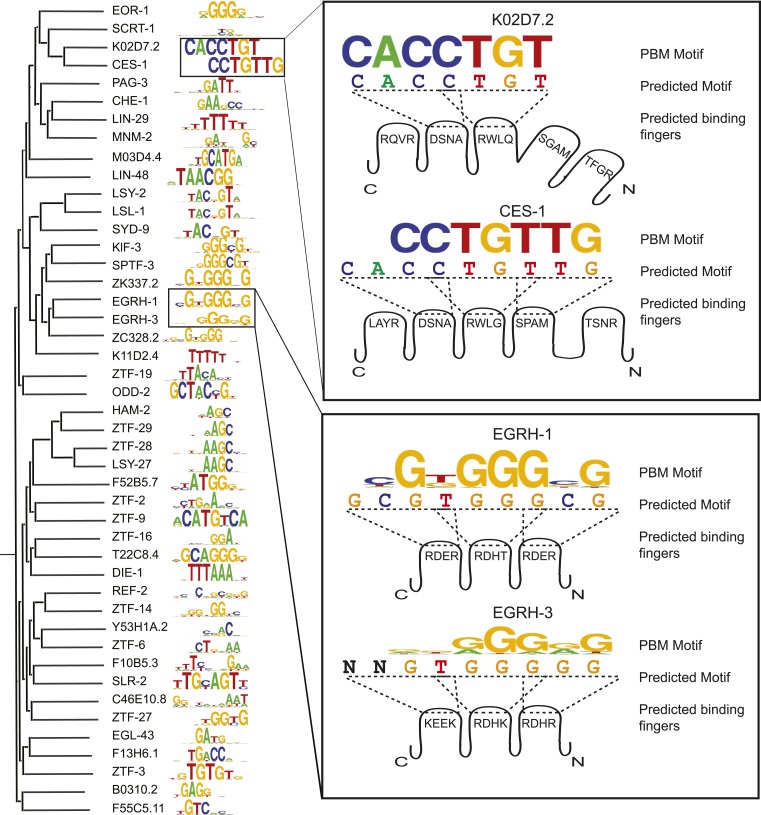
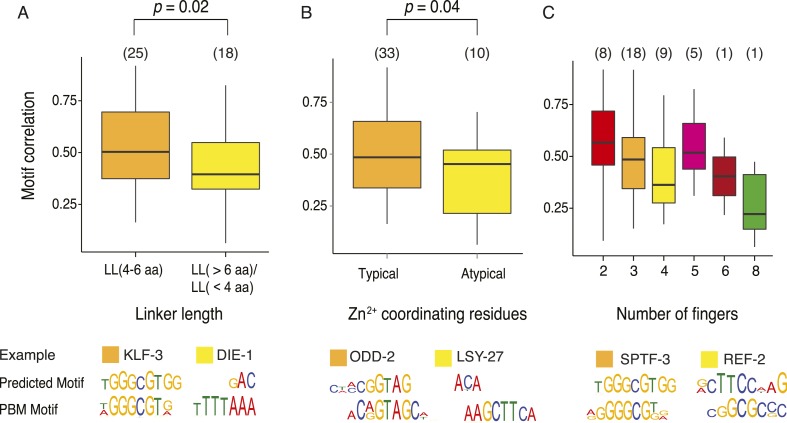
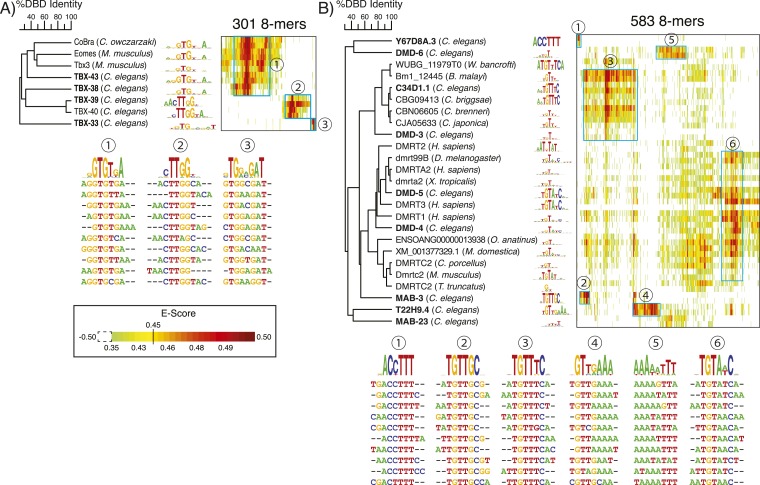
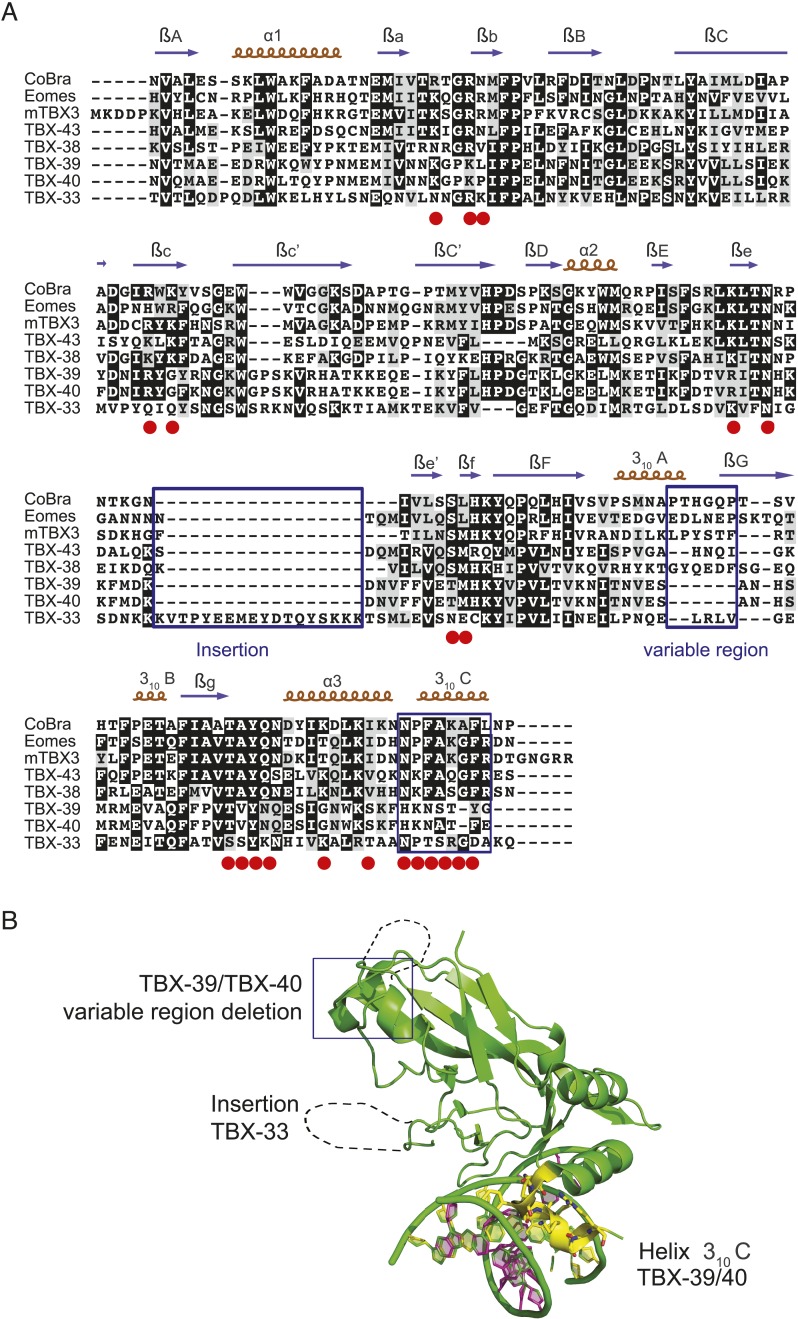
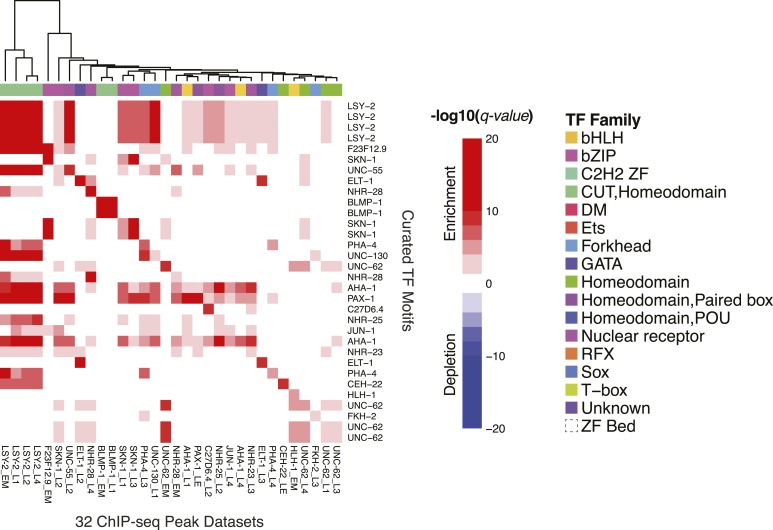
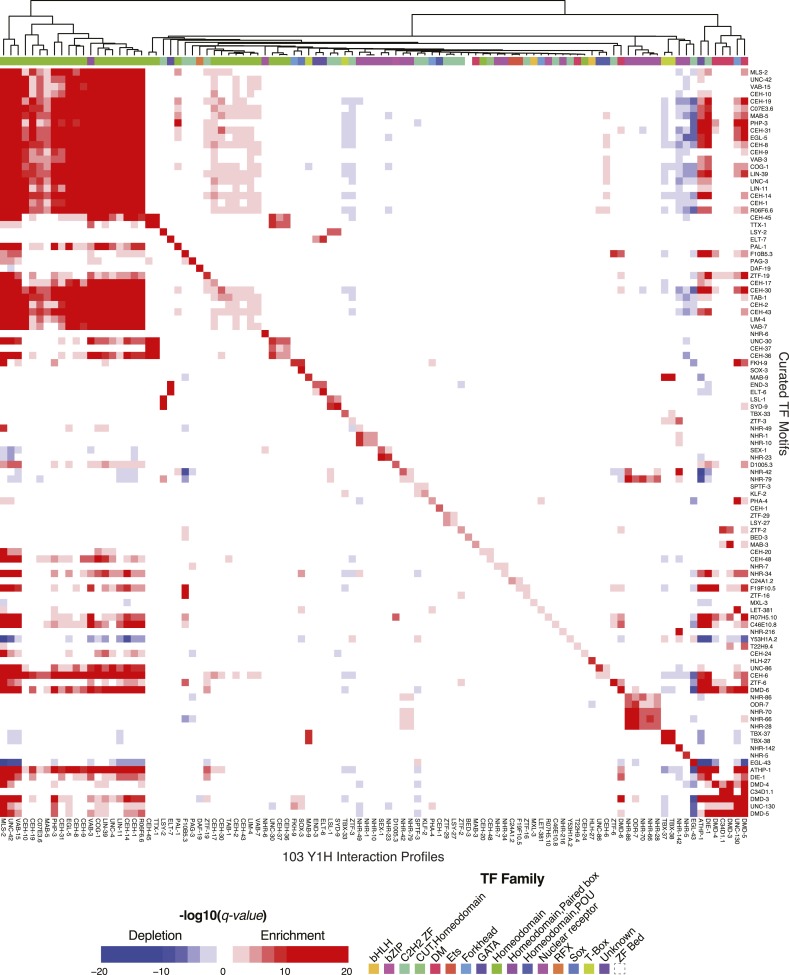
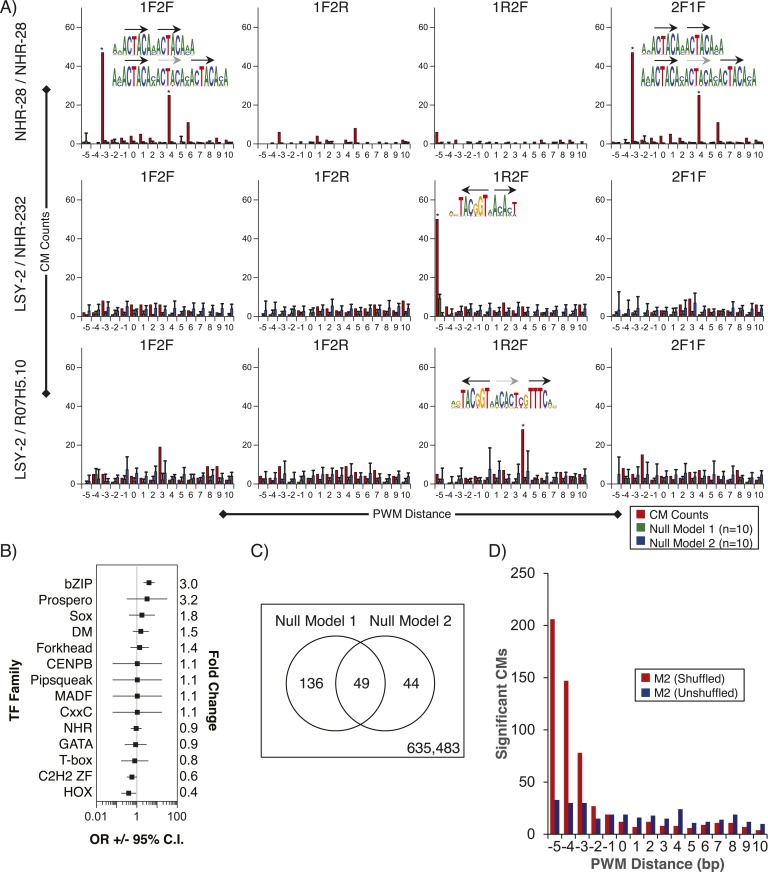
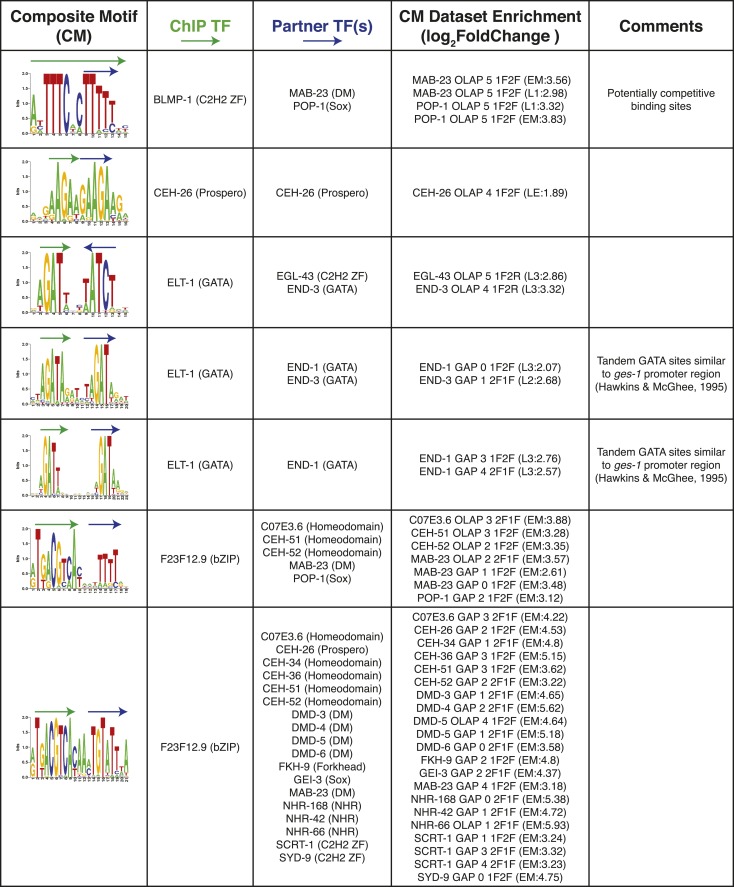
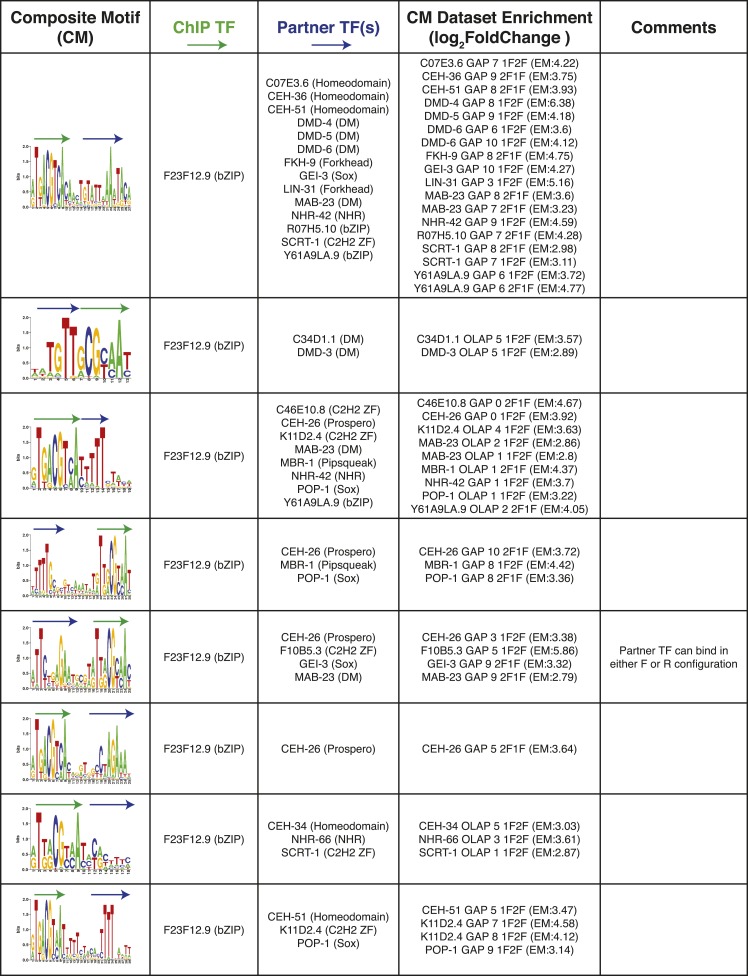
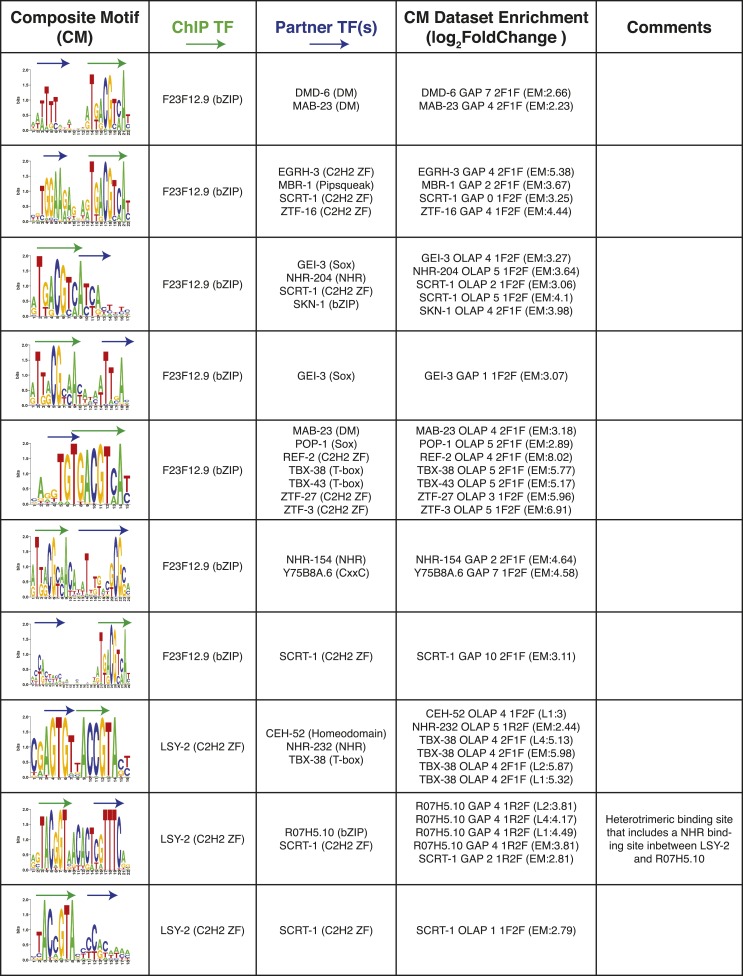
Caenorhabditis elegans is a powerful model for studying gene regulation, as it has a compact genome and a wealth of genomic tools. However, identification of regulatory elements has been limited, as DNA-binding motifs are known for only 71 of the estimated 763 sequence-specific transcription factors (TFs). To address this problem, we performed protein binding microarray experiments on representatives of canonical TF families in C. elegans, obtaining motifs for 129 TFs. Additionally, we predict motifs for many TFs that have DNA-binding domains similar to those already characterized, increasing coverage of binding specificities to 292 C. elegans TFs (∼40%). These data highlight the diversification of binding motifs for the nuclear hormone receptor and C2H2 zinc finger families and reveal unexpected diversity of motifs for T-box and DM families. Motif enrichment in promoters of functionally related genes is consistent with known biology and also identifies putative regulatory roles for unstudied TFs.
秀丽隐杆线虫是研究基因调控的强大模型,因为它具有紧凑的基因组和丰富的基因组工具。然而,调控元件的鉴定一直有限,因为在估计的763个序列特异性转录因子(TFs)中,只有71个的DNA结合基序是已知的。为了解决这个问题,我们对秀丽隐杆线虫中典型TF家族的代表进行了蛋白质结合微阵列实验,获得了129个TFs的基序。此外,我们预测了许多具有与已表征的DNA结合域相似的TFs的基序,将结合特异性的覆盖范围增加到292个秀丽隐杆线虫TFs(约40%)。这些数据突出了核激素受体和C2H2锌指家族结合基序的多样性,并揭示了T-box和DM家族基序的意外多样性。功能相关基因启动子中的基序富集与已知生物学一致,也确定了未研究TFs的假定调控作用。