Nishiya Anna Shoko, Almeida-Neto César de, Romano Camila Malta, Alencar Cecília Salete, Ferreira Suzete Cleusa, Di-Lorenzo-Oliveira Claudia, Levi José Eduardo, Salles Nanci Alves, Mendrone-Junior Alfredo, Sabino Ester Cerdeira
Fundação Pró-Sangue/Hemocentro de São Paulo, São Paulo, SP, Brazil; Infectious Diseases Division (DIPA), Universidade Federal de São Paulo (UNIFESP), São Paulo, SP, Brazil.
Fundação Pró-Sangue/Hemocentro de São Paulo, São Paulo, SP, Brazil; Discipline of Medical Science, Faculdade de Medicina, Universidade de São Paulo (USP), São Paulo, SP, Brazil.
Braz J Infect Dis. 2015 Sep-Oct;19(5):473-8. doi: 10.1016/j.bjid.2015.06.010. Epub 2015 Aug 19.
It is recognized that hepatitis C virus subtypes (1a, 1b, 2a, 2b, 2c and 3a) originated in Africa and Asia and spread worldwide exponentially during the Second World War (1940) through the transfusion of contaminated blood products, invasive medical and dental procedures, and intravenous drug use. The entry of hepatitis C virus subtypes into different regions occurred at distinct times, presenting exponential growth rates of larger or smaller spread. Our study estimated the growth and spread of the most prevalent subtypes currently circulating in São Paulo.
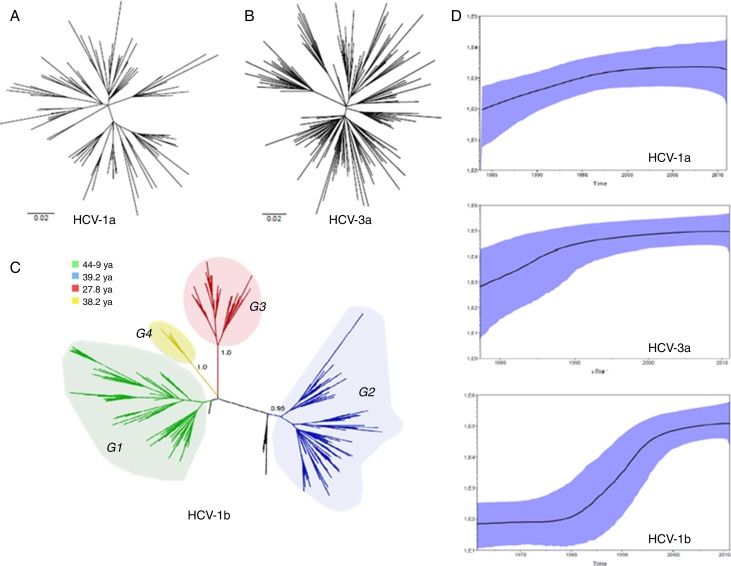
A total of 465 non-structural region 5B sequences of hepatitis C virus covering a 14-year time-span were used to reconstruct the population history and estimate the population dynamics and Time to Most Recent Common Ancestor of genotypes using the Bayesian Markov Chain Monte Carlo approach implemented in BEAST (Bayesian evolutionary analysis by sampling tree software/program).
Evolutionary analysis demonstrated that the different hepatitis C virus subtypes had distinct growth patterns. The introduction of hepatitis C virus-1a and -3a were estimated to be circa 1979 and 1967, respectively, whereas hepatitis C virus-1b appears to have a more ancient entry, circa 1923. Hepatitis C virus-1b phylogenies suggest that different lineages circulate in São Paulo, and four well-supported groups (i.e., G1, G2, G3 and G4) were identified. Hepatitis C virus-1a presented the highest growth rate (r=0.4), but its spread became less marked after the 2000s. Hepatitis C virus-3a grew exponentially until the 1990s and had an intermediate growth rate (r=0.32). An evident exponential growth (r=0.26) was found for hepatitis C virus-1b between 1980 and the mid-1990s.
After an initial period of exponential growth, the expansion of the three main subtypes began to decrease. Hepatitis C virus-1b presented inflated genetic diversity, and its transmission may have been sustained by different generations and transmission routes other than blood transfusion. Hepatitis C virus-1a and -3a showed no group stratification, most likely due to their recent entry.
人们认识到,丙型肝炎病毒亚型(1a、1b、2a、2b、2c和3a)起源于非洲和亚洲,并在第二次世界大战(1940年)期间通过受污染血液制品的输血、侵入性医疗和牙科手术以及静脉注射吸毒在全球呈指数级传播。丙型肝炎病毒亚型进入不同地区的时间各不相同,呈现出不同程度的指数增长速度。我们的研究估计了圣保罗目前流行的最普遍亚型的增长和传播情况。
总共465个涵盖14年时间跨度的丙型肝炎病毒非结构区5B序列被用于重建种群历史,并使用BEAST(通过采样树软件/程序进行贝叶斯进化分析)中实现的贝叶斯马尔可夫链蒙特卡罗方法估计种群动态和基因型的最近共同祖先时间。
进化分析表明,不同的丙型肝炎病毒亚型具有不同的增长模式。丙型肝炎病毒-1a和-3a的引入估计分别约在1979年和1967年,而丙型肝炎病毒-1b似乎有更古老的进入时间,约在1923年左右。丙型肝炎病毒-1b系统发育表明,不同的谱系在圣保罗传播,并且鉴定出了四个得到充分支持的组(即G1、G2、G3和G4)。丙型肝炎病毒-1a呈现出最高的增长率(r = 0.4),但其在21世纪后传播变得不那么明显。丙型肝炎病毒-3a在20世纪90年代之前呈指数增长,并且具有中等增长率(r = 0.32)。在1980年至20世纪90年代中期之间,丙型肝炎病毒-1b出现了明显的指数增长(r = 0.26)。
在最初的指数增长期之后,三种主要亚型的扩张开始减少。丙型肝炎病毒-1b呈现出膨胀的遗传多样性,并且其传播可能通过不同代以及输血以外的其他传播途径得以持续。丙型肝炎病毒-1a和-3a没有显示出群体分层,最有可能是由于它们近期才进入。