Martins Nuno M C, Bergmann Jan H, Shono Nobuaki, Kimura Hiroshi, Larionov Vladimir, Masumoto Hiroshi, Earnshaw William C
Wellcome Trust Centre for Cell Biology, University of Edinburgh, Edinburgh EH9 3JR, Scotland, United Kingdom.
Laboratory of Cell Engineering, Department of Frontier Research, Kazusa DNA Research Institute, Kisarazu 292-0818, Japan Division of Biological Science, Graduate School of Science, Nagoya University, Nagoya 464-8602, Japan.
Mol Biol Cell. 2016 Jan 1;27(1):177-96. doi: 10.1091/mbc.E15-08-0605. Epub 2015 Nov 12.
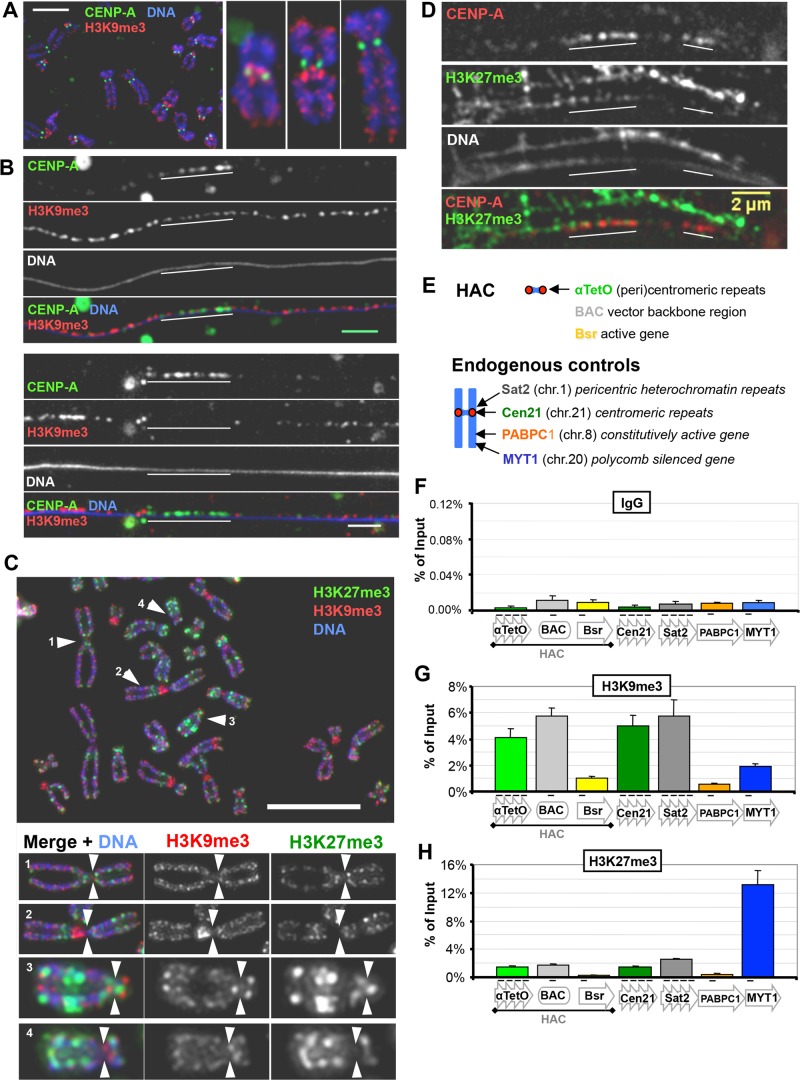
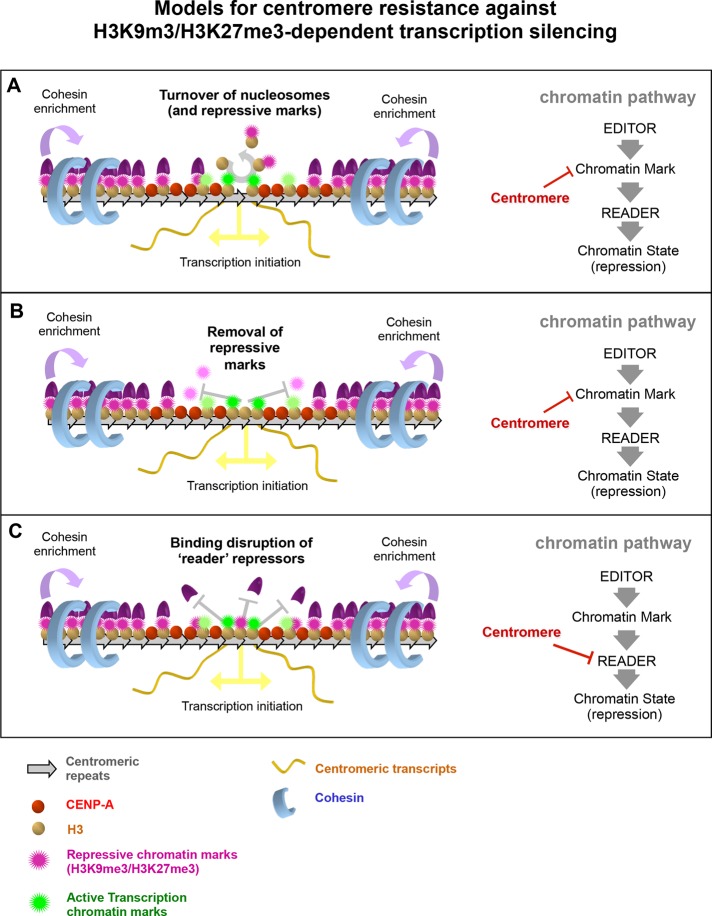
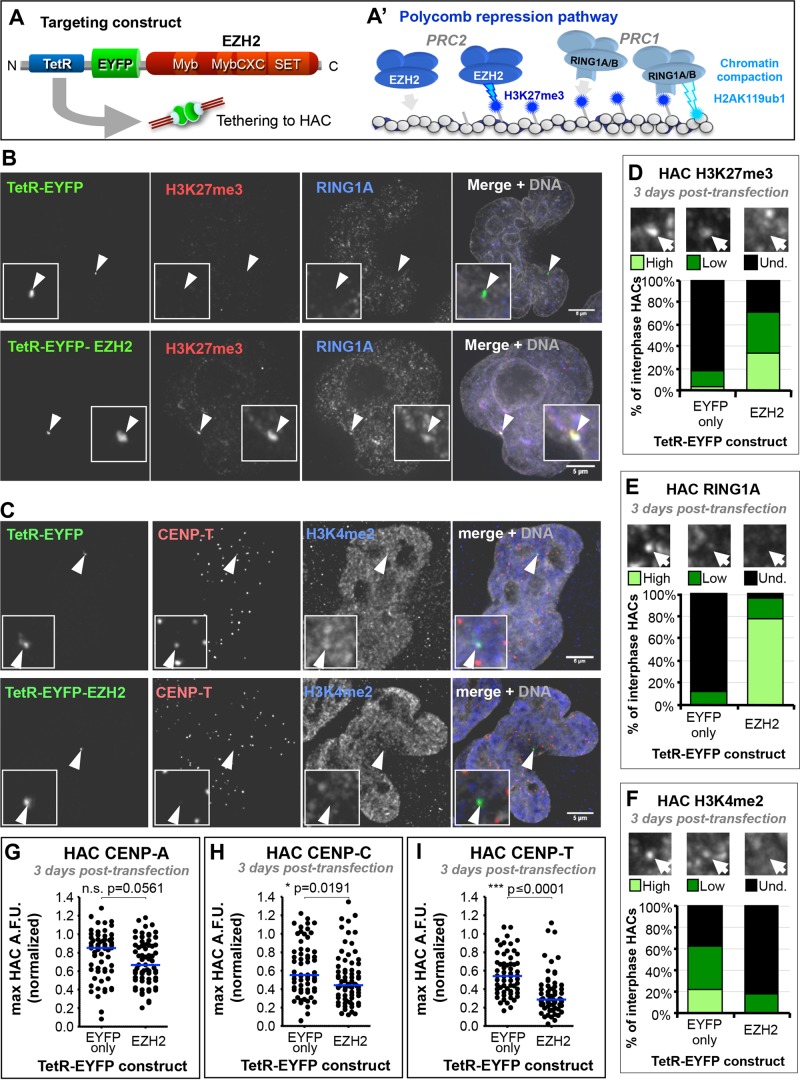
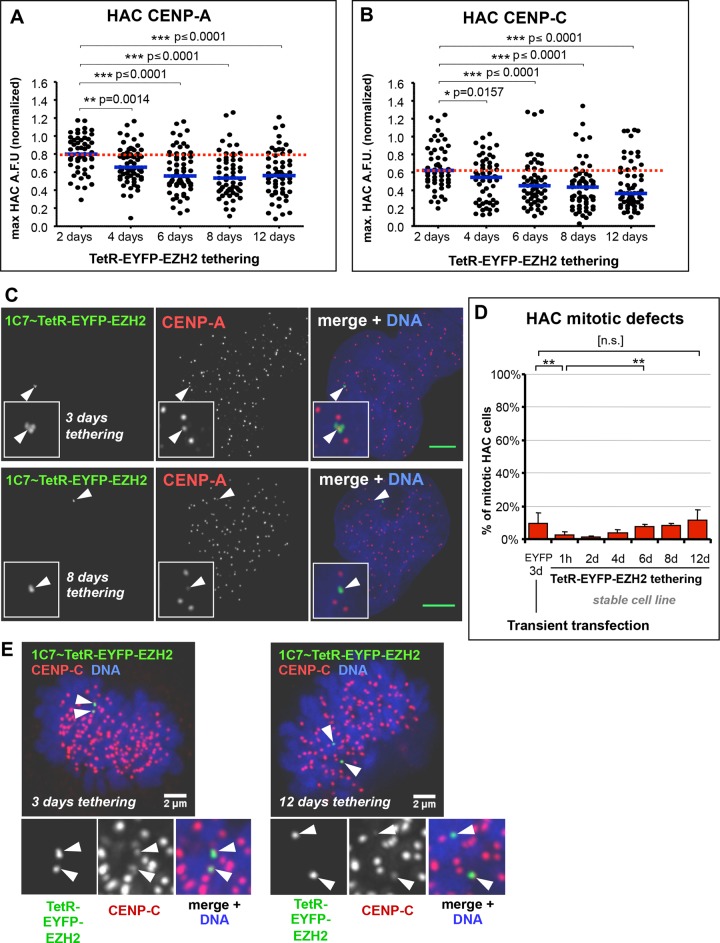
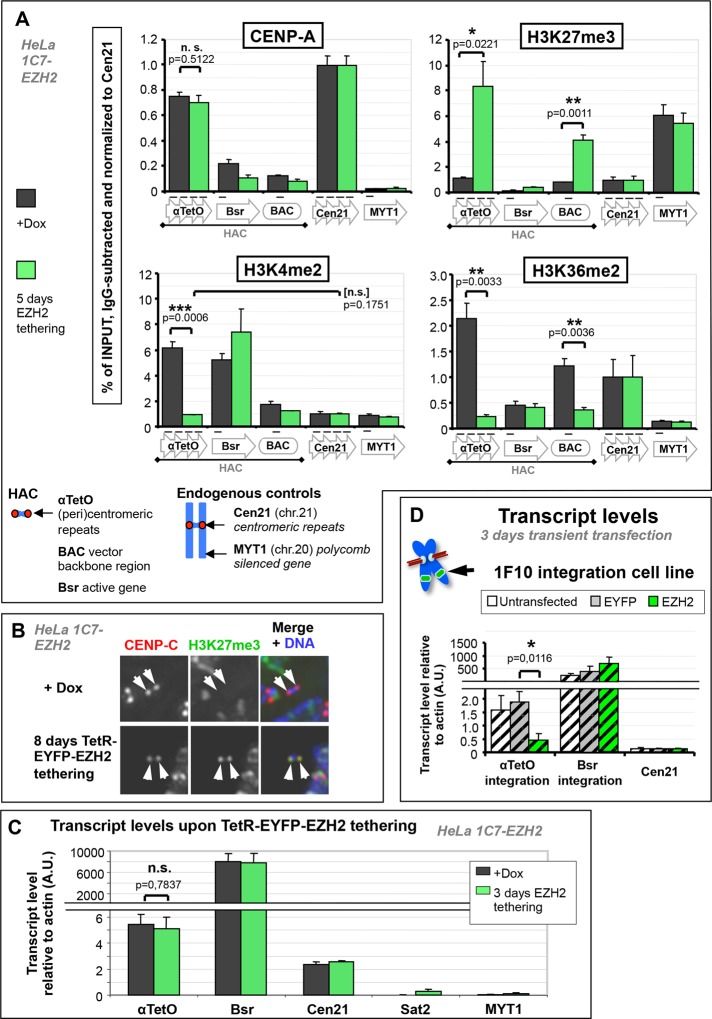
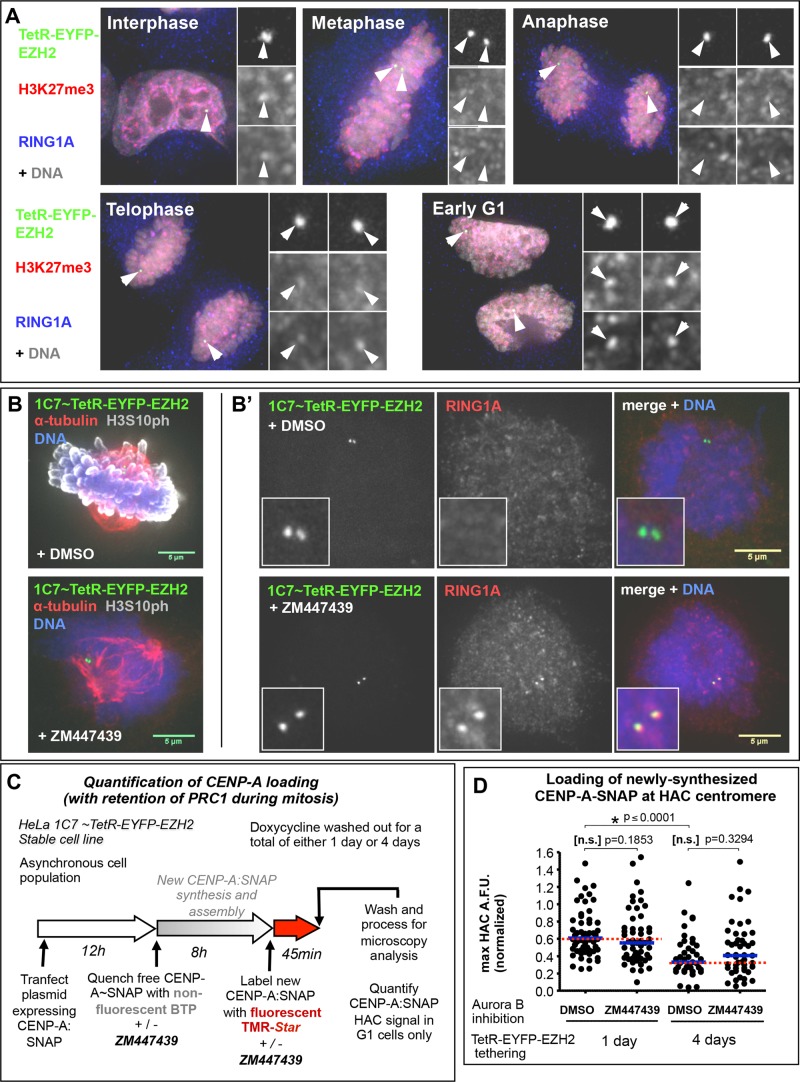
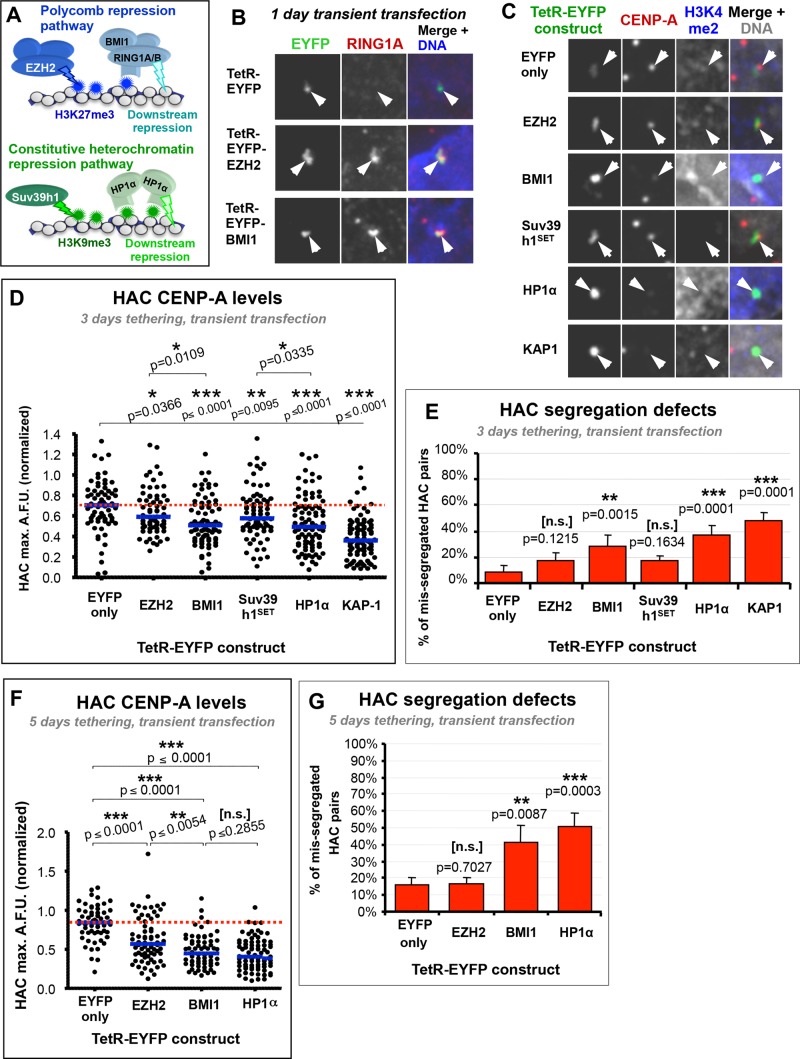
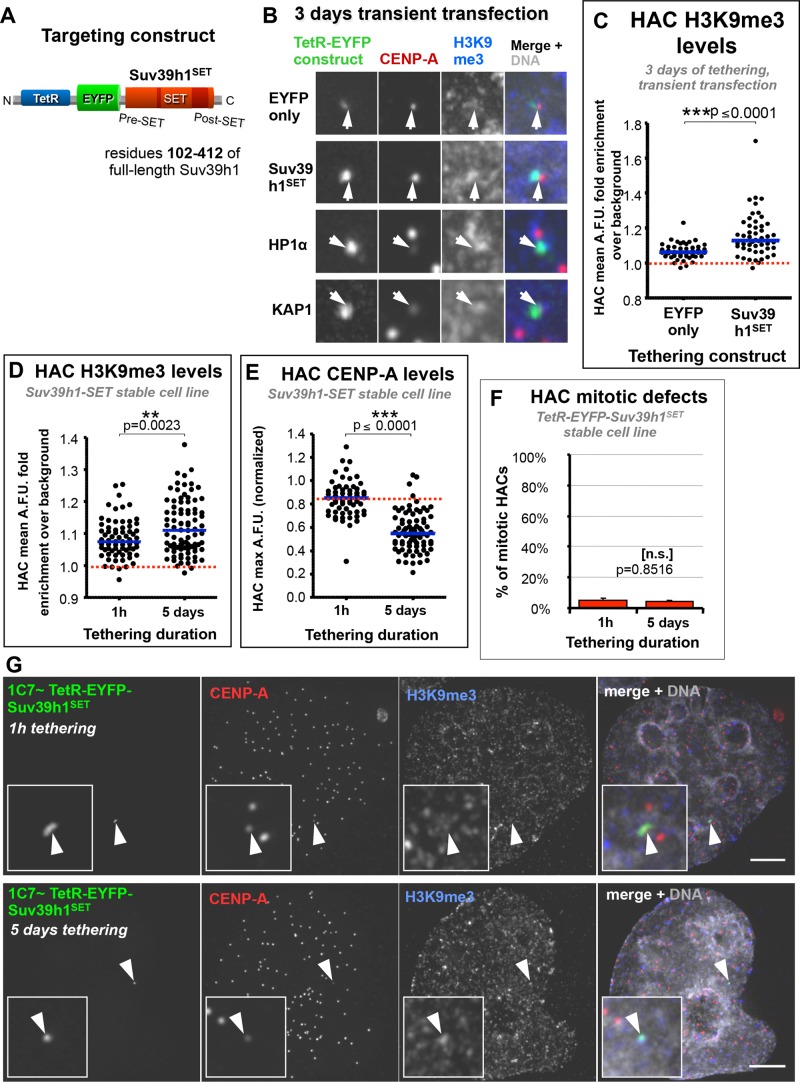
Centromeres are characterized by the centromere-specific H3 variant CENP-A, which is embedded in chromatin with a pattern characteristic of active transcription that is required for centromere identity. It is unclear how centromeres remain transcriptionally active despite being flanked by repressive pericentric heterochromatin. To further understand centrochromatin's response to repressive signals, we nucleated a Polycomb-like chromatin state within the centromere of a human artificial chromosome (HAC) by tethering the methyltransferase EZH2. This led to deposition of the H3K27me3 mark and PRC1 repressor binding. Surprisingly, this state did not abolish HAC centromere function or transcription, and this apparent resistance was not observed on a noncentromeric locus, where transcription was silenced. Directly tethering the reader/repressor PRC1 bypassed this resistance, inactivating the centromere. We observed analogous responses when tethering the heterochromatin Editor Suv39h1-methyltransferase domain (centromere resistance) or reader HP1α (centromere inactivation), respectively. Our results reveal that the HAC centromere can resist repressive pathways driven by H3K9me3/H3K27me3 and may help to explain how centromeres are able to resist inactivation by flanking heterochromatin.
着丝粒的特征是含有着丝粒特异性的组蛋白H3变体CENP-A,它嵌入具有活跃转录特征模式的染色质中,这对于着丝粒的特性是必需的。目前尚不清楚着丝粒如何在被抑制性的着丝粒周围异染色质包围的情况下仍保持转录活性。为了进一步了解着丝粒染色质对抑制信号的反应,我们通过拴系甲基转移酶EZH2在人类人工染色体(HAC)的着丝粒内诱导了一种类似多梳蛋白的染色质状态。这导致了H3K27me3标记的沉积和PRC1阻遏物的结合。令人惊讶的是,这种状态并没有消除HAC着丝粒的功能或转录,并且在转录被沉默的非着丝粒位点上没有观察到这种明显的抗性。直接拴系读取器/阻遏物PRC1绕过了这种抗性,使着丝粒失活。当分别拴系异染色质编辑器Suv39h1甲基转移酶结构域(着丝粒抗性)或读取器HP1α(着丝粒失活)时,我们观察到了类似的反应。我们的结果表明,HAC着丝粒可以抵抗由H3K9me3/H3K27me3驱动的抑制途径,这可能有助于解释着丝粒如何能够抵抗侧翼异染色质的失活作用。