Garcia Benjamin A, Luka Zigmund, Loukachevitch Lioudmila V, Bhanu Natarajan V, Wagner Conrad
Epigenetics Program, Departments of Biochemistry and Biophysics, Perelman School of Medicine, University of Pennsylvania, 3400 Civic Center Blvd, Philadelphia, PA 19104, USA.
Department of Biochemistry, Vanderbilt University School of Medicine, 21st Avenue South, Nashville, TN 37232, USA.
Med Hypotheses. 2016 Mar;88:63-7. doi: 10.1016/j.mehy.2015.12.027. Epub 2016 Jan 9.
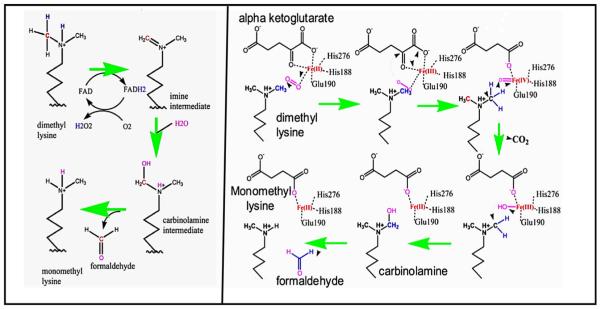
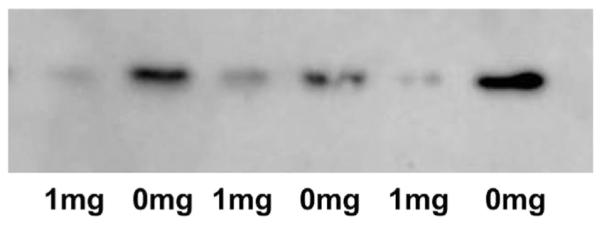
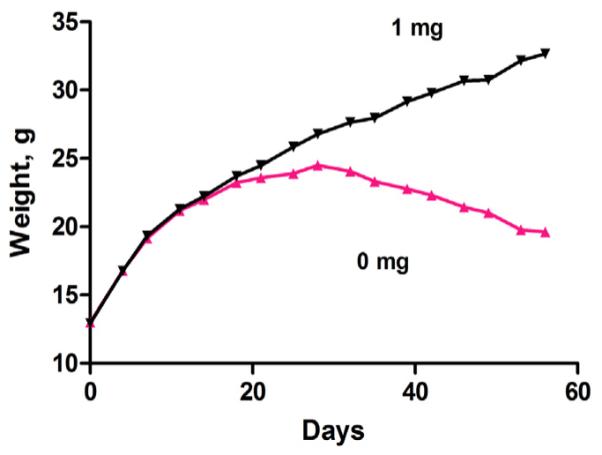
Formaldehyde is extremely toxic reacting with proteins to crosslinks peptide chains. Formaldehyde is a metabolic product in many enzymatic reactions and the question of how these enzymes are protected from the formaldehyde that is generated has largely remained unanswered. Early experiments from our laboratory showed that two liver mitochondrial enzymes, dimethylglycine dehydrogenase (DMGDH) and sarcosine dehydrogenase (SDH) catalyze oxidative demethylation reactions (sarcosine is a common name for monomethylglycine). The enzymatic products of these enzymes were the demethylated substrates and formaldehyde, produced from the removed methyl group. Both DMGDH and SDH contain FAD and both have tightly bound tetrahydrofolate (THF), a folate coenzyme. THF binds reversibly with formaldehyde to form 5,10-methylene-THF. At that time we showed that purified DMGDH, with tightly bound THF, reacted with formaldehyde generated during the reaction to form 5,10-methylene-THF. This effectively scavenged the formaldehyde to protect the enzyme. Recently, post-translational modifications on histone tails have been shown to be responsible for epigenetic regulation of gene expression. One of these modifications is methylation of lysine residues. The first enzyme discovered to accomplish demethylation of these modified histones was histone lysine demethylase (LSD1). LSD1 specifically removes methyl groups from di- and mono-methylated lysines at position 4 of histone 3. This enzyme contained tightly bound FAD and the products of the reaction were the demethylated lysine residue and formaldehyde. The mechanism of LSD1 demethylation is analogous to the mechanism previously postulated for DMGDH, i.e. oxidation of the N-methyl bond to the methylene imine followed by hydrolysis to generate formaldehyde. This suggested that THF might also be involved in the LSD1 reaction to scavenge the formaldehyde produced. Our hypotheses are that THF is bound to native LSD1 by analogy to DMGDH and SDH and that the bound THF serves to protect the FAD class of histone demethylases from the destructive effects of formaldehyde generation by formation of 5,10-methylene-THF. We present pilot data showing that decreased folate in livers as a result of dietary folate deficiency is associated with increased levels of methylated lysine 4 of histone 3. This can be a result of decreased LSD1 activity resulting from the decreased folate available to scavenge the formaldehyde produced at the active site caused by the folate deficiency. Because LSD1 can regulate gene expression this suggests that folate may play a more important role than simply serving as a carrier of one-carbon units and be a factor in other diseases associated with low folate.
甲醛具有极高的毒性,它会与蛋白质发生反应,使肽链交联。甲醛是许多酶促反应的代谢产物,而这些酶如何免受所生成甲醛的影响这一问题在很大程度上仍未得到解答。我们实验室早期的实验表明,两种肝脏线粒体酶,即二甲基甘氨酸脱氢酶(DMGDH)和肌氨酸脱氢酶(SDH),催化氧化脱甲基反应(肌氨酸是一甲基甘氨酸的常用名称)。这些酶的酶促产物是脱甲基化的底物和由去除的甲基产生的甲醛。DMGDH和SDH都含有黄素腺嘌呤二核苷酸(FAD),并且都紧密结合有四氢叶酸(THF),一种叶酸辅酶。THF与甲醛可逆结合形成5,10 - 亚甲基 - THF。当时我们表明,紧密结合有THF的纯化DMGDH会与反应过程中生成的甲醛反应形成5,10 - 亚甲基 - THF。这有效地清除了甲醛以保护该酶。最近,已表明组蛋白尾部的翻译后修饰负责基因表达的表观遗传调控。这些修饰之一是赖氨酸残基的甲基化。发现的第一种完成这些修饰组蛋白脱甲基化的酶是组蛋白赖氨酸去甲基化酶(LSD1)。LSD1特异性地从组蛋白3第4位的二甲基化和单甲基化赖氨酸上去除甲基。这种酶紧密结合有FAD,反应产物是脱甲基化的赖氨酸残基和甲醛。LSD1脱甲基化的机制类似于先前为DMGDH假设的机制,即N - 甲基键氧化为亚甲基亚胺,然后水解生成甲醛。这表明THF也可能参与LSD1反应以清除产生的甲醛。我们的假设是,类似于DMGDH和SDH,THF与天然LSD1结合,并且结合的THF通过形成5,10 - 亚甲基 - THF来保护FAD类组蛋白去甲基化酶免受甲醛生成的破坏作用。我们提供的初步数据表明,由于饮食中叶酸缺乏导致肝脏中叶酸减少与组蛋白3赖氨酸4甲基化水平升高有关。这可能是由于叶酸缺乏导致活性位点产生的甲醛可用于清除的叶酸减少,从而导致LSD1活性降低的结果。因为LSD1可以调节基因表达,这表明叶酸可能发挥比仅仅作为一碳单位载体更重要的作用,并且是与低叶酸相关的其他疾病的一个因素。