Kollimada Somanna A, Kulkarni Ankur H, Ravan Aniket, Gundiah Namrata
Department of Mechanical Engineering, Indian Institute of Science, Bangalore, India.
Department of Molecular Reproduction, Development and Genetics, Indian Institute of Science, Bangalore, India.
PLoS One. 2016 Apr 14;11(4):e0153471. doi: 10.1371/journal.pone.0153471. eCollection 2016.
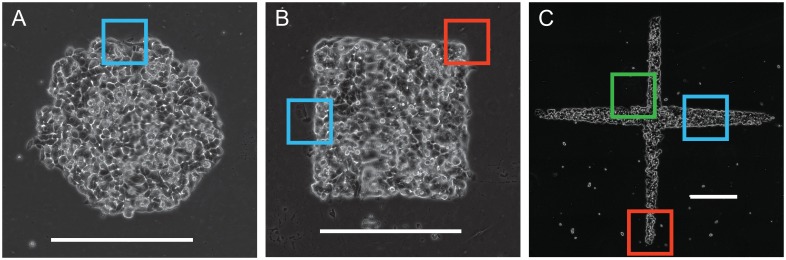
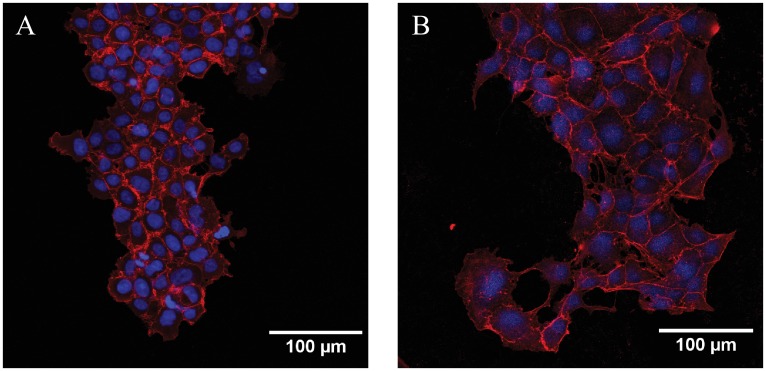
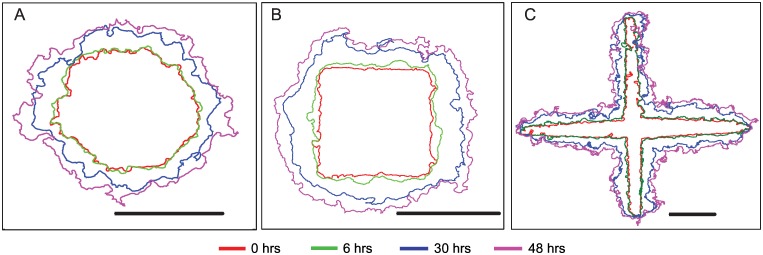
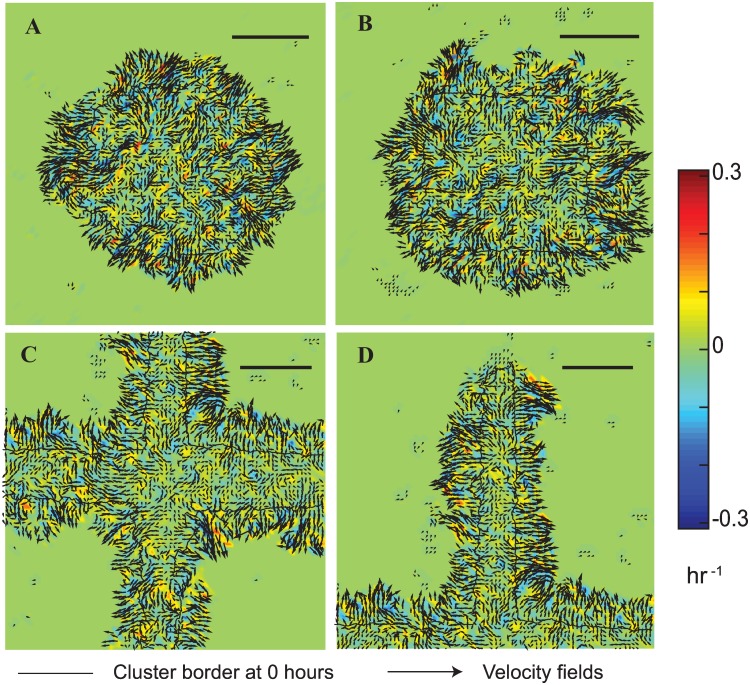
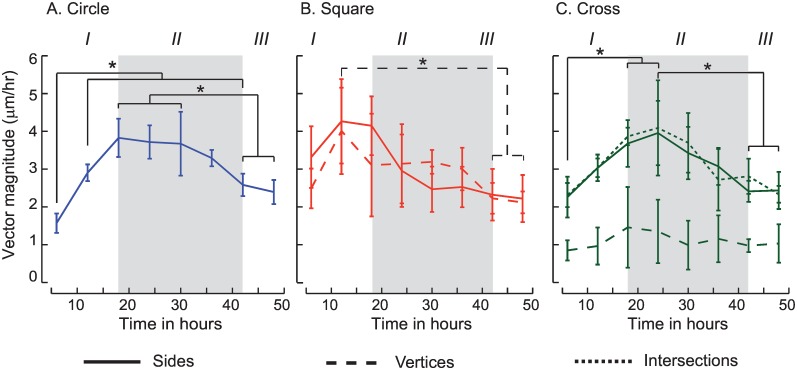
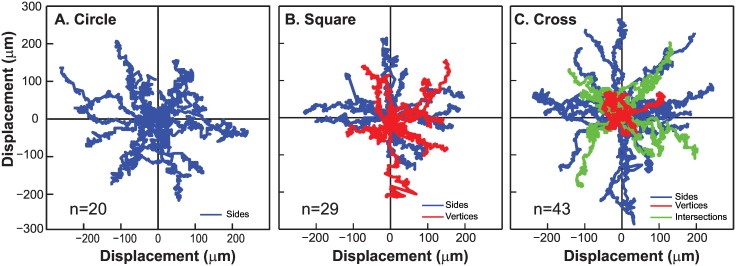
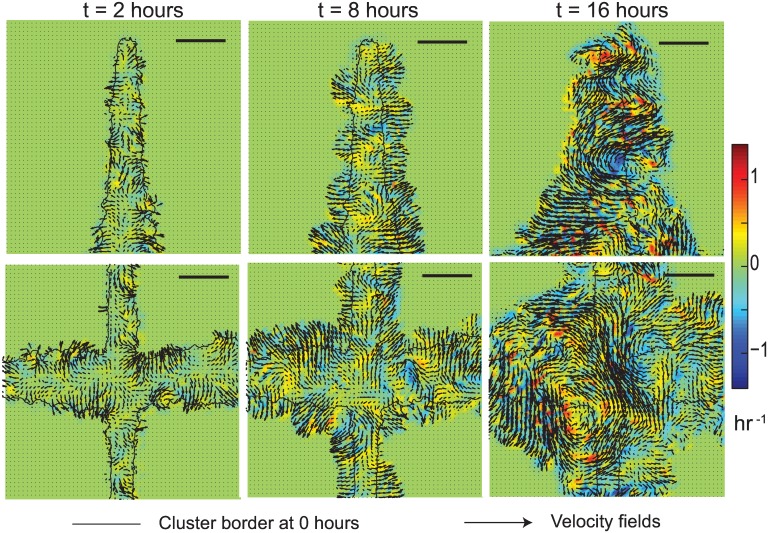
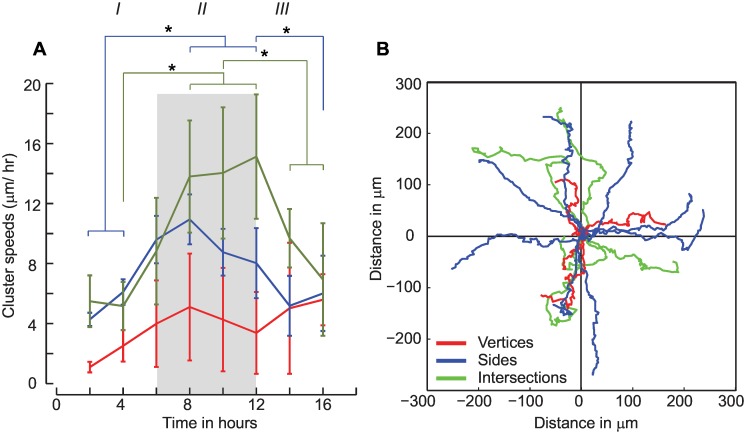
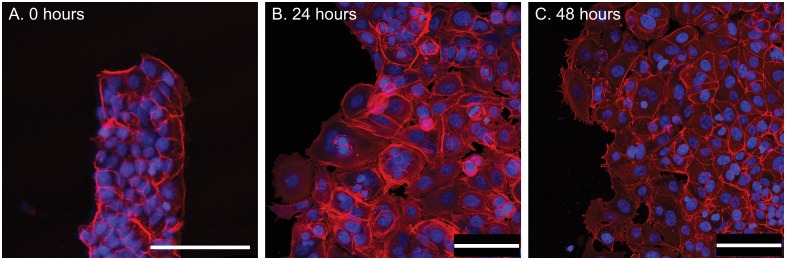
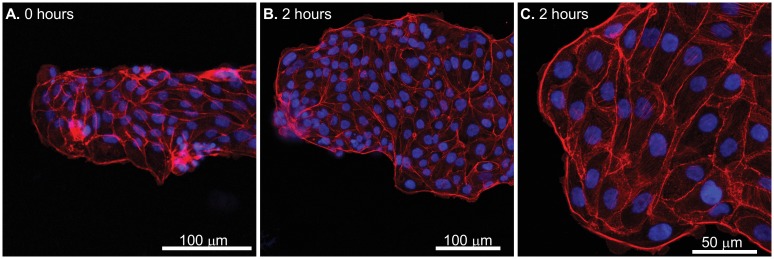
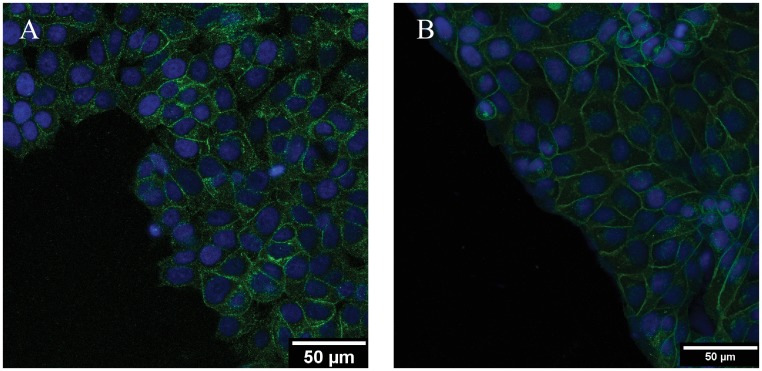
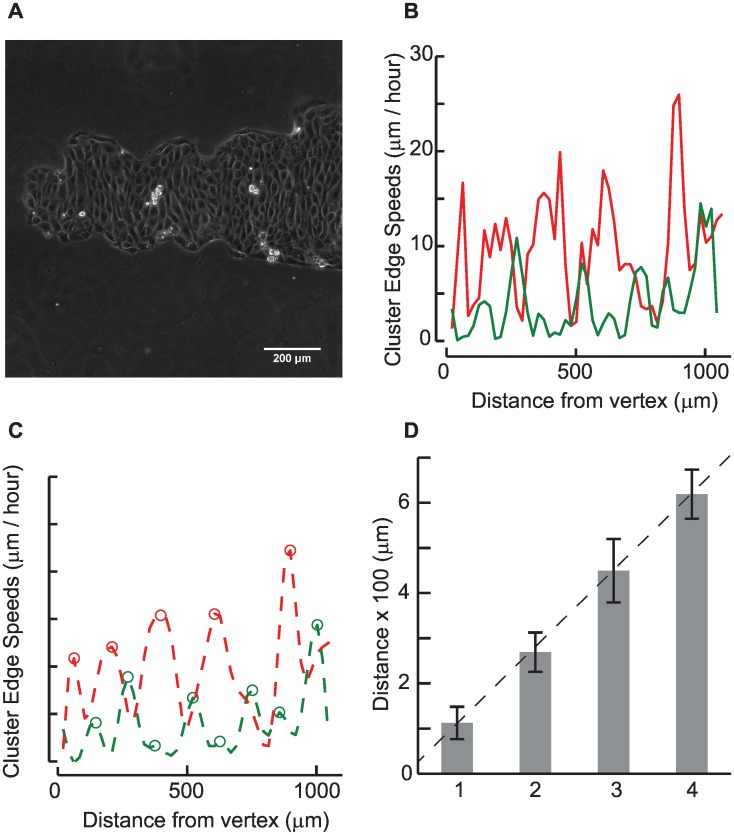
Collective cell migrations are essential in several physiological processes and are driven by both chemical and mechanical cues. The roles of substrate stiffness and confinement on collective migrations have been investigated in recent years, however few studies have addressed how geometric shapes influence collective cell migrations. Here, we address the hypothesis that the relative position of a cell within the confinement influences its motility. Monolayers of two types of epithelial cells--MCF7, a breast epithelial cancer cell line, and MDCK, a control epithelial cell line--were confined within circular, square, and cross-shaped stencils and their migration velocities were quantified upon release of the constraint using particle image velocimetry. The choice of stencil geometry allowed us to investigate individual cell motility within convex, straight and concave boundaries. Cells located in sharp, convex boundaries migrated at slower rates than those in concave or straight edges in both cell types. The overall cluster migration occurred in three phases: an initial linear increase with time, followed by a plateau region and a subsequent decrease in cluster speeds. An acto-myosin contractile ring, present in the MDCK but absent in MCF7 monolayer, was a prominent feature in the emergence of leader cells from the MDCK clusters which occurred every ~125 μm from the vertex of the cross. Further, coordinated cell movements displayed vorticity patterns in MDCK which were absent in MCF7 clusters. We also used cytoskeletal inhibitors to show the importance of acto-myosin bounding cables in collective migrations through translation of local movements to create long range coordinated movements and the creation of leader cells within ensembles. To our knowledge, this is the first demonstration of how bounding shapes influence long-term migratory behaviours of epithelial cell monolayers. These results are important for tissue engineering and may also enhance our understanding of cell movements during developmental patterning and cancer metastasis.
集体细胞迁移在多个生理过程中至关重要,且由化学和机械信号驱动。近年来,人们研究了底物硬度和限制条件对集体迁移的作用,然而,很少有研究探讨几何形状如何影响集体细胞迁移。在此,我们探讨这样一个假设:细胞在限制条件内的相对位置会影响其运动性。将两种上皮细胞单层——乳腺癌上皮细胞系MCF7和对照上皮细胞系MDCK——限制在圆形、方形和十字形模板内,并在解除限制后使用粒子图像测速法对其迁移速度进行量化。模板几何形状的选择使我们能够研究凸形、直线形和凹形边界内单个细胞的运动性。在两种细胞类型中,位于尖锐凸形边界的细胞迁移速度均低于凹形或直线形边界的细胞。整体集群迁移分为三个阶段:最初随时间呈线性增加,随后是平稳期,集群速度随后下降。在MDCK单层中存在而在MCF7单层中不存在的肌动蛋白-肌球蛋白收缩环,是MDCK集群中每隔约125μm从十字顶点出现的领头细胞的一个显著特征。此外,MDCK中的协调细胞运动显示出涡旋模式,而MCF7集群中则没有。我们还使用细胞骨架抑制剂来证明肌动蛋白-肌球蛋白结合索在集体迁移中的重要性,即通过局部运动的平移来产生长距离协调运动,并在群体中产生领头细胞。据我们所知,这是首次证明边界形状如何影响上皮细胞单层的长期迁移行为。这些结果对组织工程很重要,也可能增进我们对发育模式形成和癌症转移过程中细胞运动的理解。