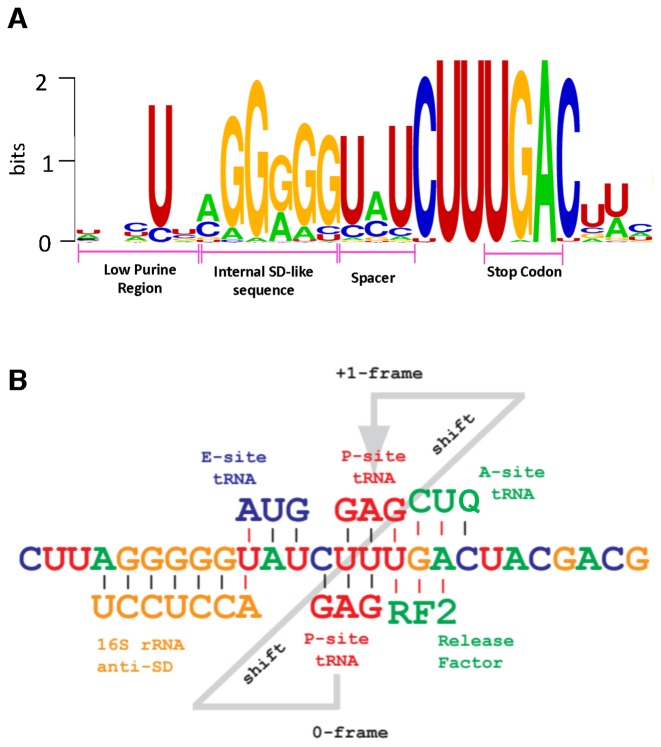
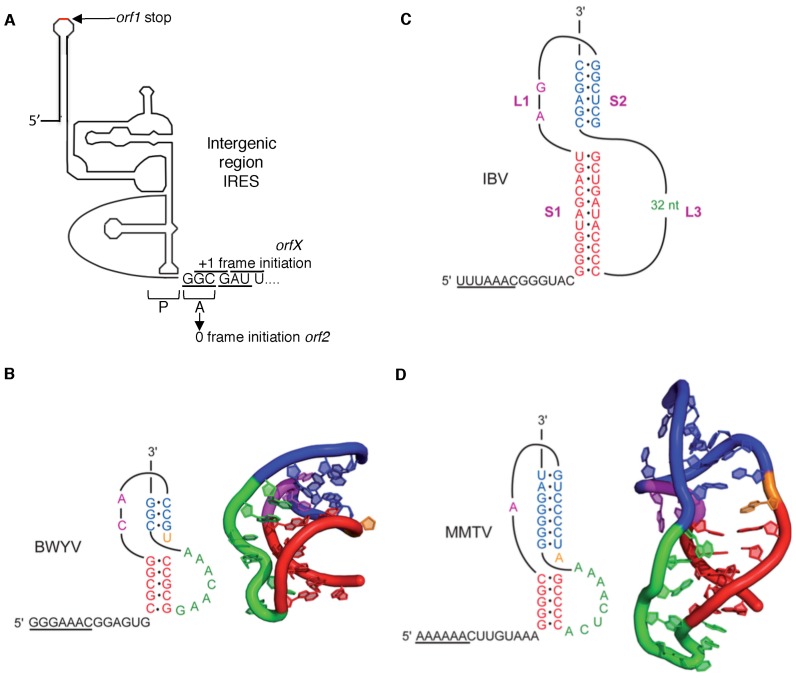
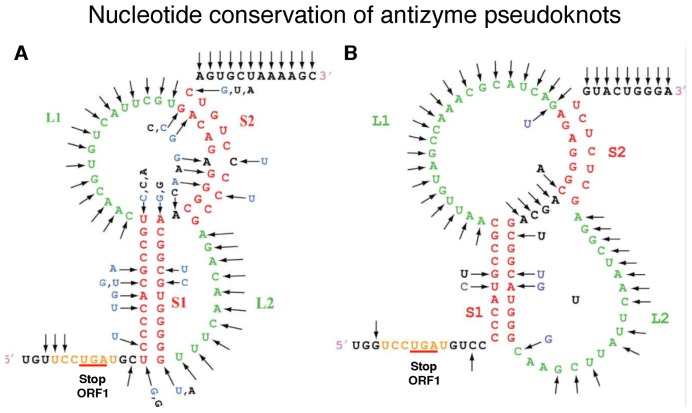
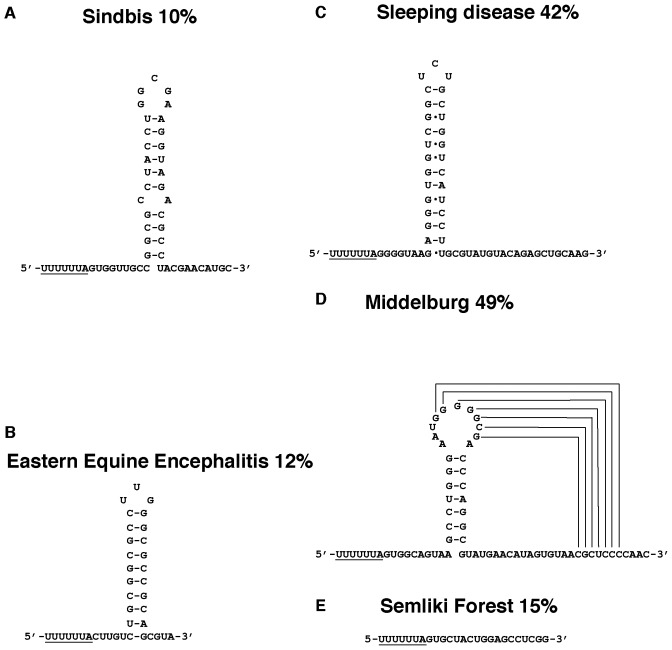
Atkins John F, Loughran Gary, Bhatt Pramod R, Firth Andrew E, Baranov Pavel V
School of Biochemistry and Cell Biology, University College Cork, Cork, Ireland School of Microbiology, University College Cork, Cork, Ireland Department of Human Genetics, University of Utah, Salt Lake City, UT 84112, USA
School of Biochemistry and Cell Biology, University College Cork, Cork, Ireland.
Nucleic Acids Res. 2016 Sep 6;44(15):7007-78. doi: 10.1093/nar/gkw530. Epub 2016 Jul 19.
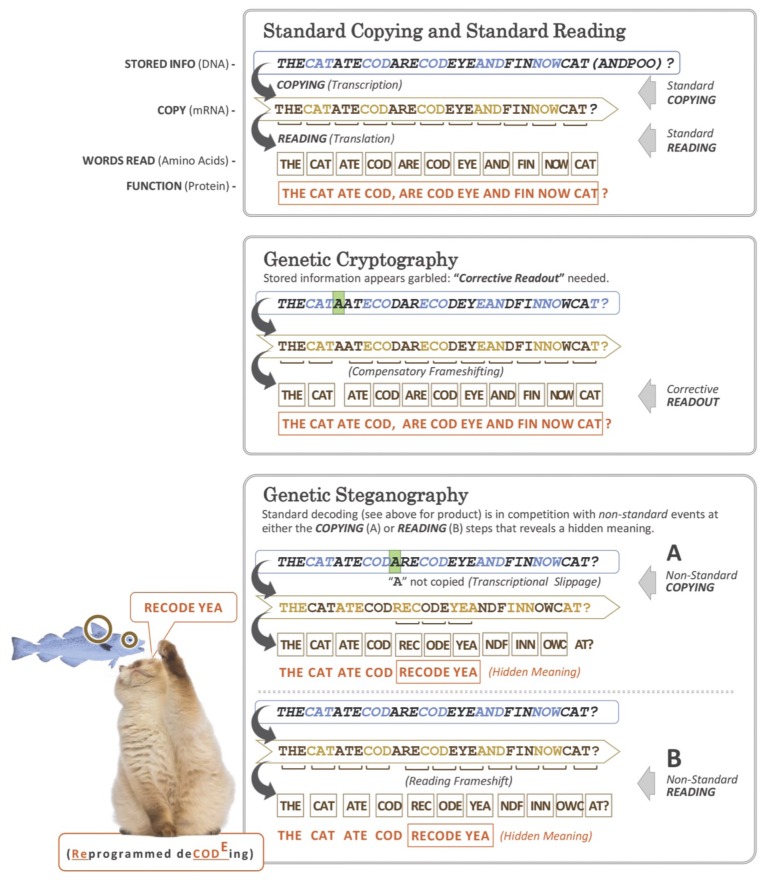
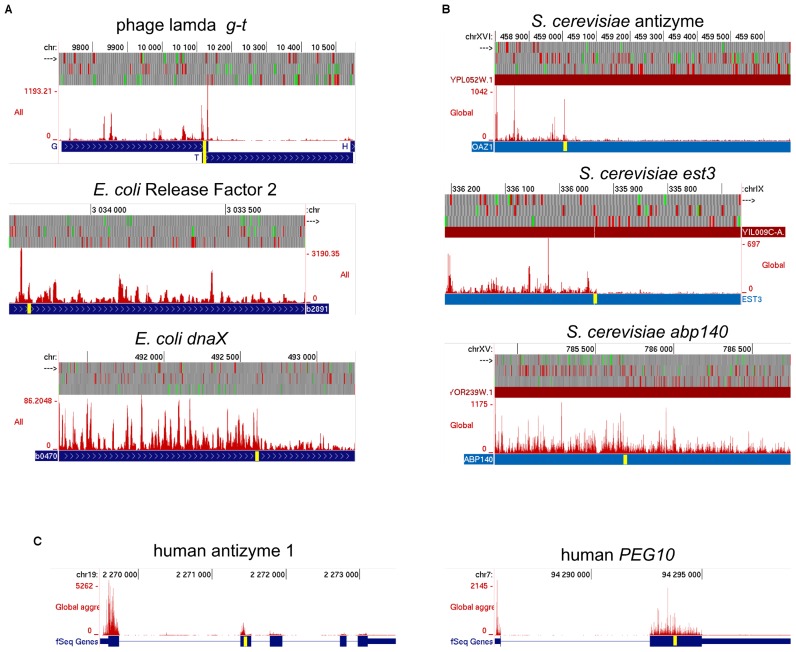
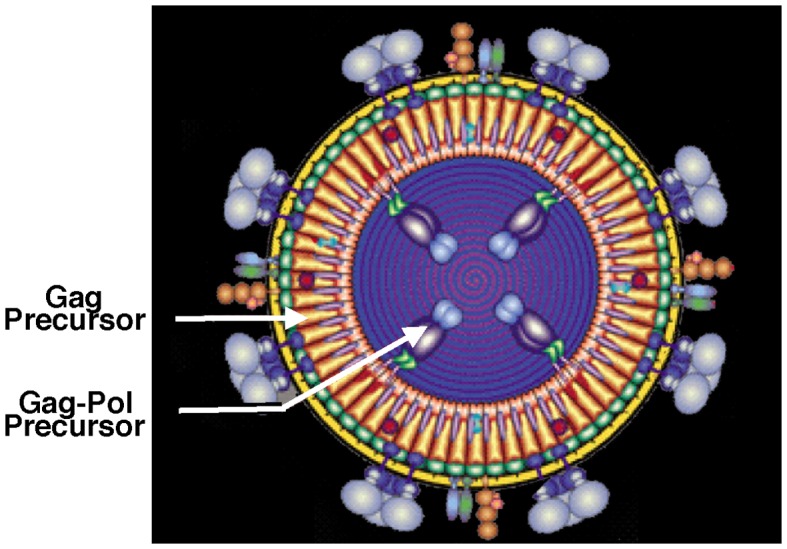
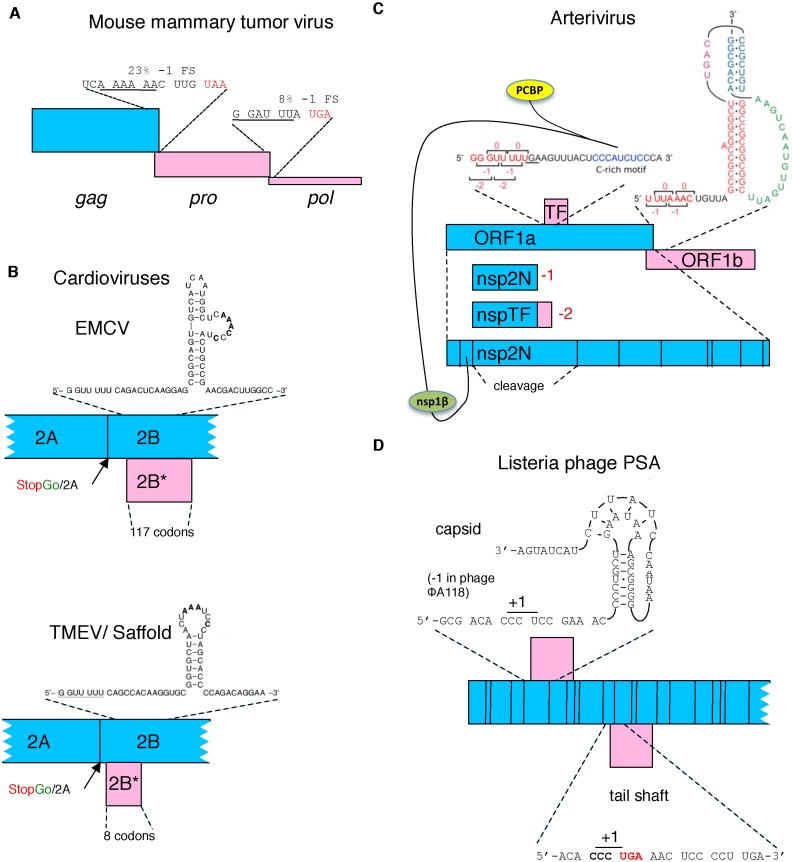
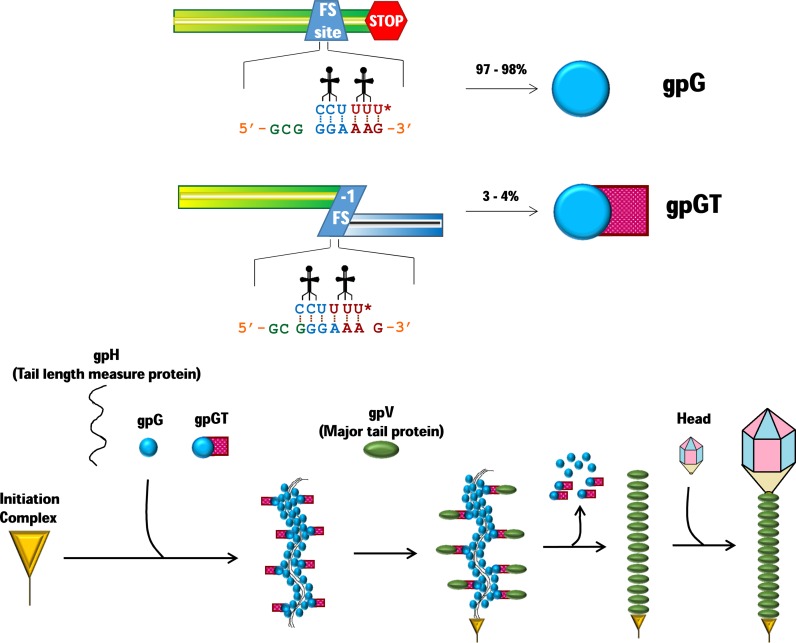
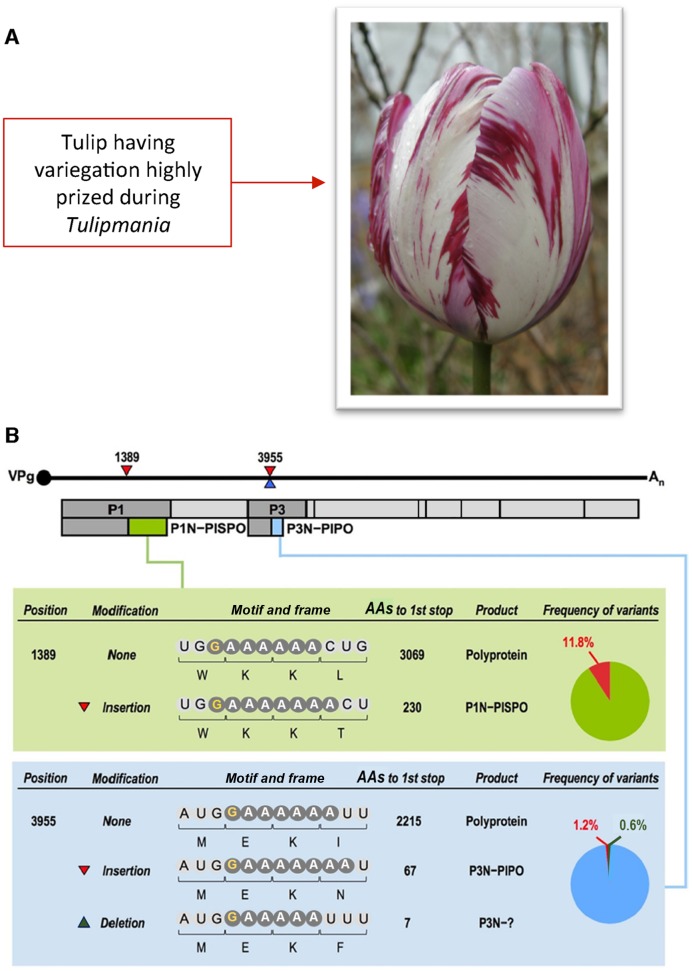
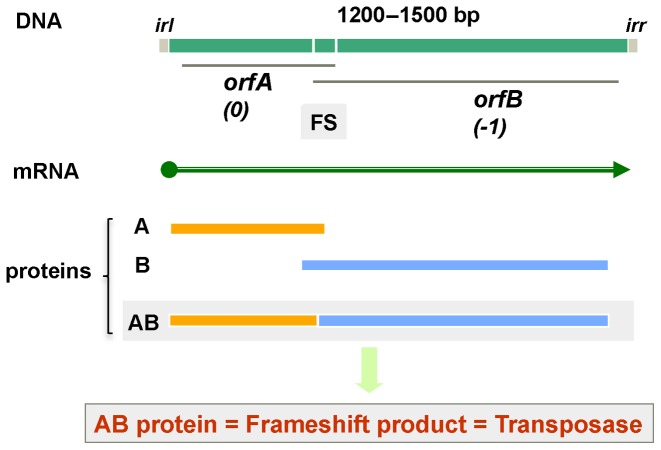
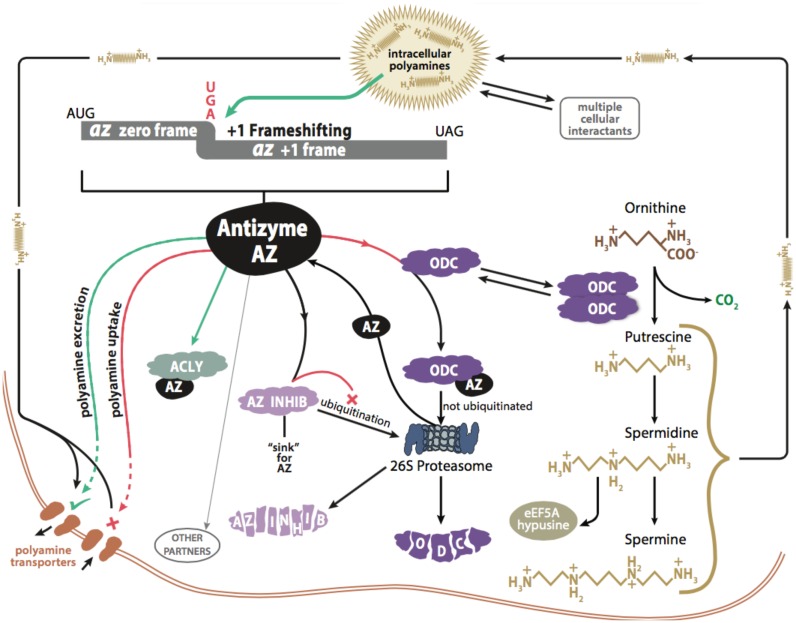
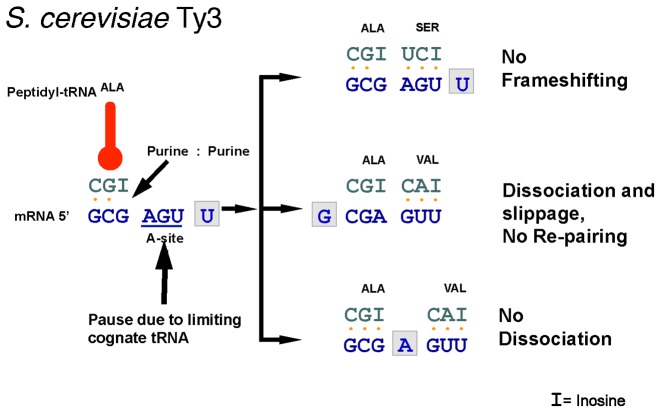
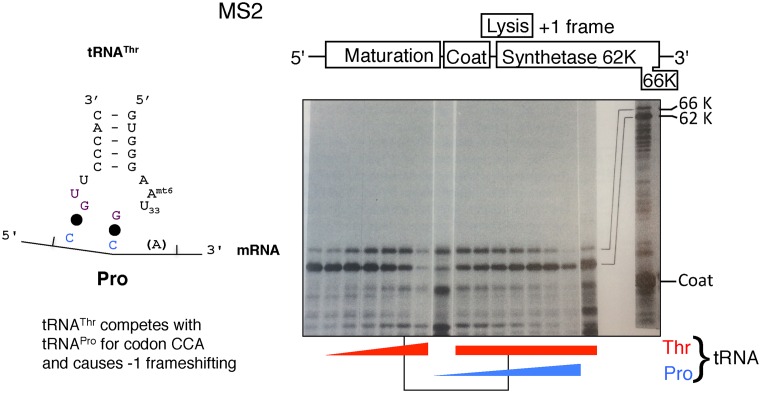
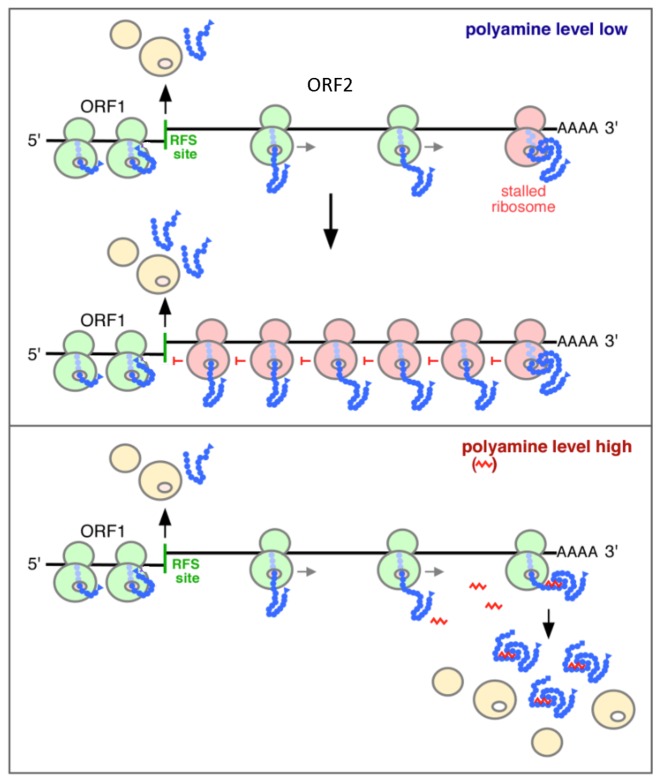
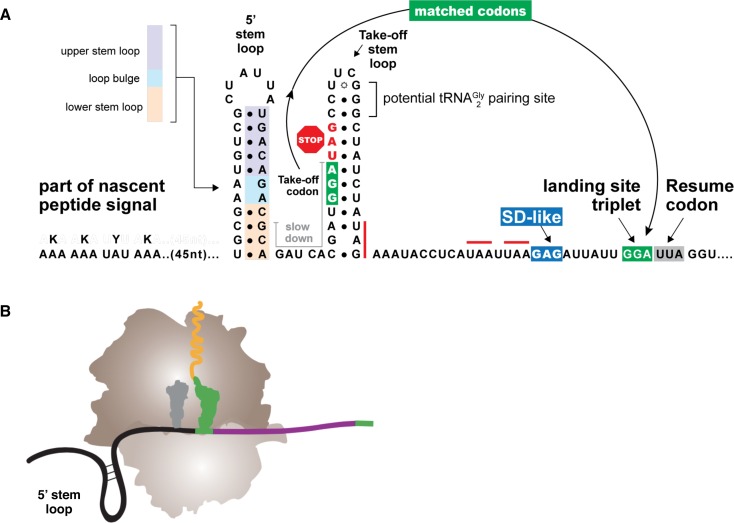
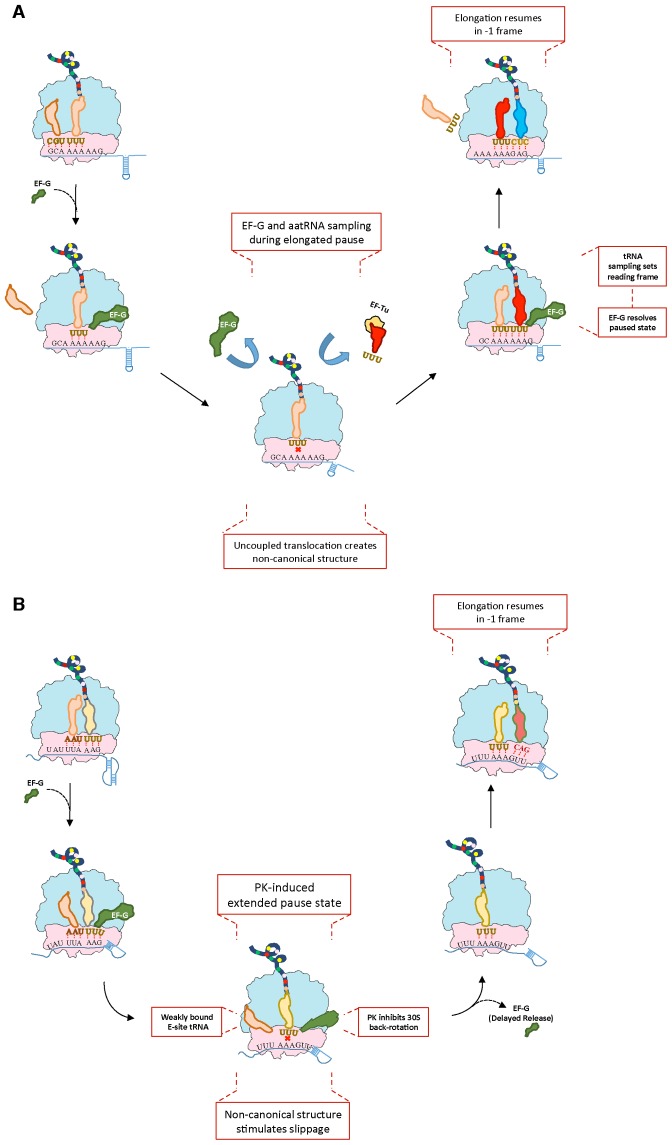
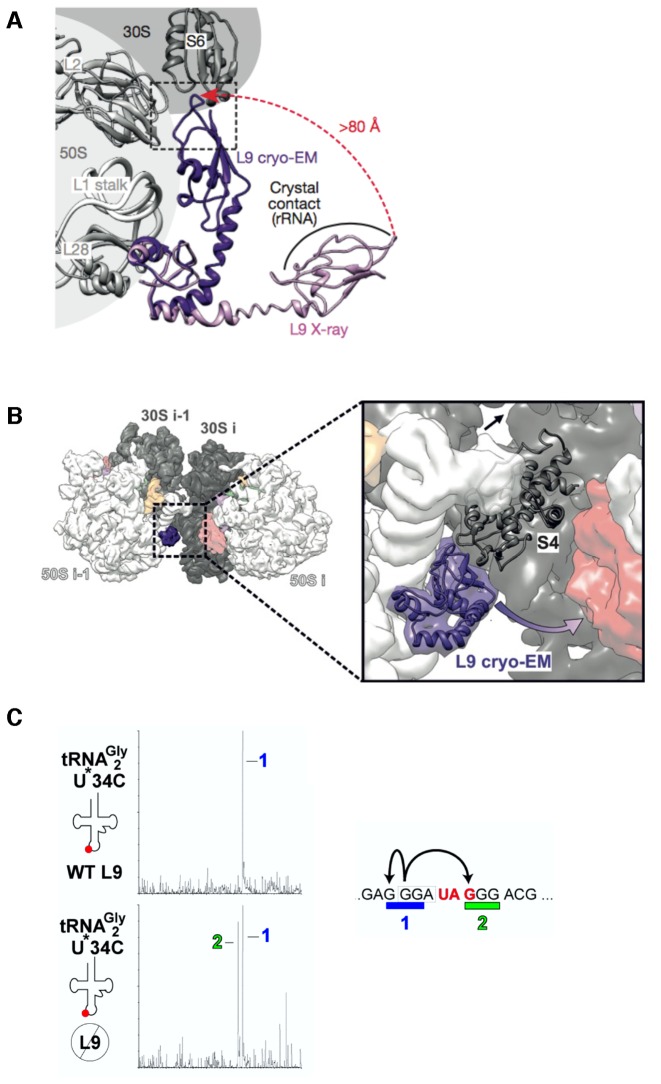
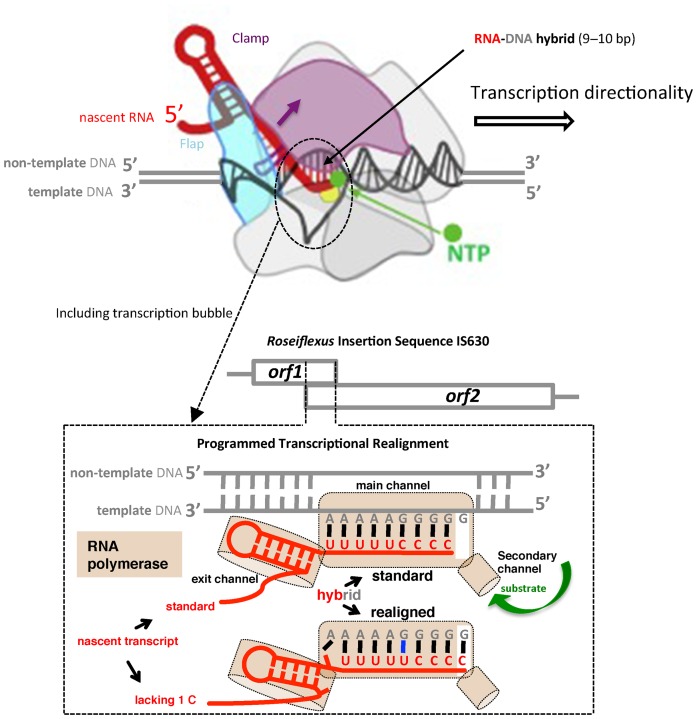
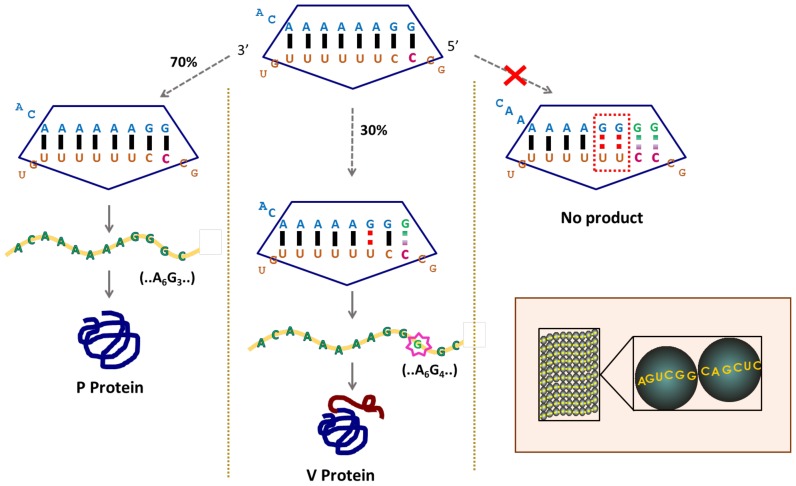
Genetic decoding is not 'frozen' as was earlier thought, but dynamic. One facet of this is frameshifting that often results in synthesis of a C-terminal region encoded by a new frame. Ribosomal frameshifting is utilized for the synthesis of additional products, for regulatory purposes and for translational 'correction' of problem or 'savior' indels. Utilization for synthesis of additional products occurs prominently in the decoding of mobile chromosomal element and viral genomes. One class of regulatory frameshifting of stable chromosomal genes governs cellular polyamine levels from yeasts to humans. In many cases of productively utilized frameshifting, the proportion of ribosomes that frameshift at a shift-prone site is enhanced by specific nascent peptide or mRNA context features. Such mRNA signals, which can be 5' or 3' of the shift site or both, can act by pairing with ribosomal RNA or as stem loops or pseudoknots even with one component being 4 kb 3' from the shift site. Transcriptional realignment at slippage-prone sequences also generates productively utilized products encoded trans-frame with respect to the genomic sequence. This too can be enhanced by nucleic acid structure. Together with dynamic codon redefinition, frameshifting is one of the forms of recoding that enriches gene expression.
基因解码并非如之前所认为的那样是“固定不变的”,而是动态的。其中一个方面是移码,这通常会导致由新的读码框编码的C末端区域的合成。核糖体移码被用于合成额外的产物、用于调控目的以及对问题插入缺失或“挽救性”插入缺失进行翻译“校正”。用于合成额外产物在移动染色体元件和病毒基因组的解码中尤为突出。稳定染色体基因的一类调控性移码控制着从酵母到人类的细胞多胺水平。在许多有效利用移码的情况下,在易发生移码的位点发生移码的核糖体比例会因特定的新生肽或mRNA上下文特征而增加。这样的mRNA信号,可以在移码位点的5'端或3'端,或者两端都有,它可以通过与核糖体RNA配对起作用,或者作为茎环或假结起作用,甚至其中一个元件在离移码位点3'端4 kb处。在易发生滑动的序列处的转录重排也会产生相对于基因组序列以反式读码框编码的有效利用的产物。这同样可以通过核酸结构得到增强。与动态密码子重新定义一起,移码是丰富基因表达的重新编码形式之一。