Abley Katie, Sauret-Güeto Susanna, Marée Athanasius Fm, Coen Enrico
John Innes Centre, Norwich Research Park, Norwich, United Kingdom.
Elife. 2016 Aug 1;5:e18165. doi: 10.7554/eLife.18165.
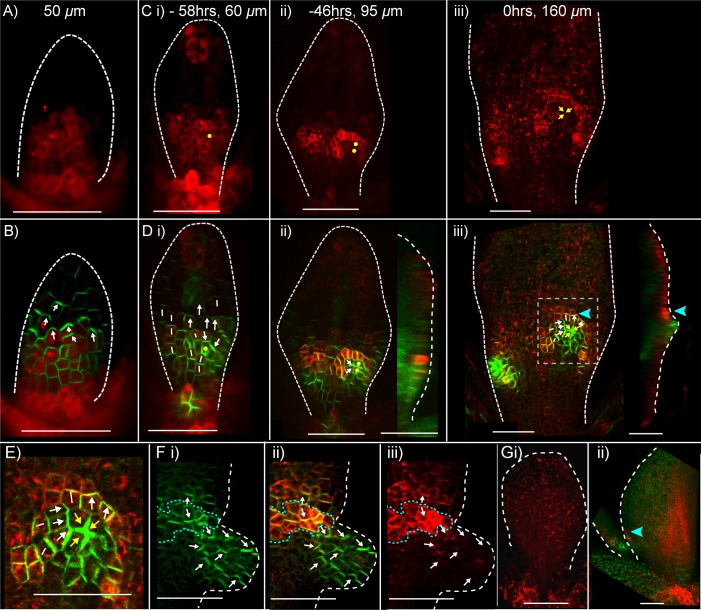
The development of outgrowths from plant shoots depends on formation of epidermal sites of cell polarity convergence with high intracellular auxin at their centre. A parsimonious model for generation of convergence sites is that cell polarity for the auxin transporter PIN1 orients up auxin gradients, as this spontaneously generates convergent alignments. Here we test predictions of this and other models for the patterns of auxin biosynthesis and import. Live imaging of outgrowths from kanadi1 kanadi2 Arabidopsis mutant leaves shows that they arise by formation of PIN1 convergence sites within a proximodistal polarity field. PIN1 polarities are oriented away from regions of high auxin biosynthesis enzyme expression, and towards regions of high auxin importer expression. Both expression patterns are required for normal outgrowth emergence, and may form part of a common module underlying shoot outgrowths. These findings are more consistent with models that spontaneously generate tandem rather than convergent alignments.
植物茎尖长出物的发育取决于在其中心具有高细胞内生长素的细胞极性汇聚的表皮位点的形成。生成汇聚位点的一个简约模型是,生长素转运蛋白PIN1的细胞极性沿生长素梯度向上定向,因为这会自发产生汇聚排列。在这里,我们测试了这个模型以及其他关于生长素生物合成和输入模式的模型的预测。对拟南芥kanadi1 kanadi2突变体叶片长出物的实时成像显示,它们是通过在近远轴极性场内形成PIN1汇聚位点而产生的。PIN1极性远离生长素生物合成酶高表达区域,而朝向生长素输入体高表达区域。这两种表达模式都是正常长出物出现所必需的,并且可能构成茎尖长出物潜在共同模块的一部分。这些发现与自发产生串联而非汇聚排列的模型更为一致。