van Kruijsbergen Ila, Hontelez Saartje, Elurbe Dei M, van Heeringen Simon J, Huynen Martijn A, Veenstra Gert Jan C
Radboud University, Department of Molecular Developmental Biology, Radboud Institute for Molecular Life Sciences, Faculty of Science, PO Box 9101, 6500 HB Nijmegen, The Netherlands.
Radboud University Medical Center, Center for Molecular and Biomolecular Informatics, Radboud Institute for Molecular Life Sciences, PO Box 9101, 6500 HB Nijmegen, The Netherlands.
Dev Biol. 2017 Jun 15;426(2):460-471. doi: 10.1016/j.ydbio.2016.08.031. Epub 2016 Sep 14.
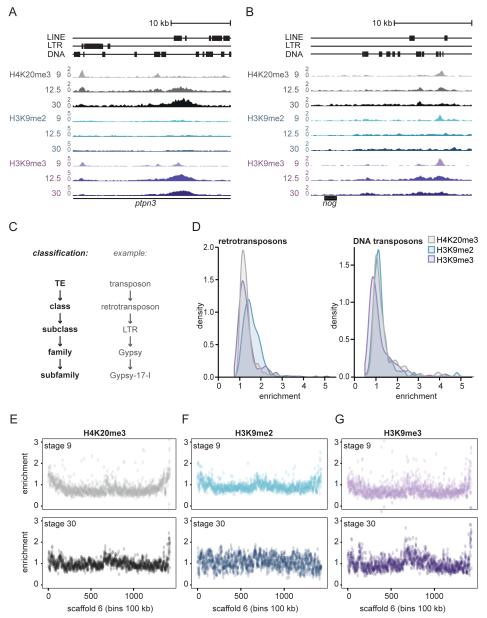
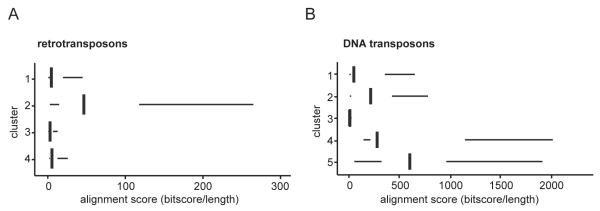
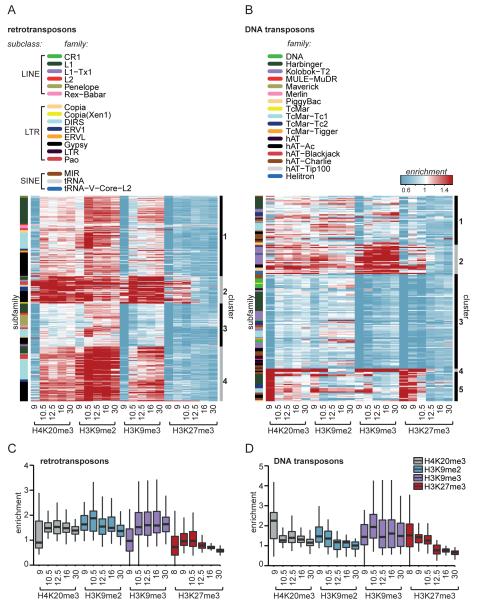
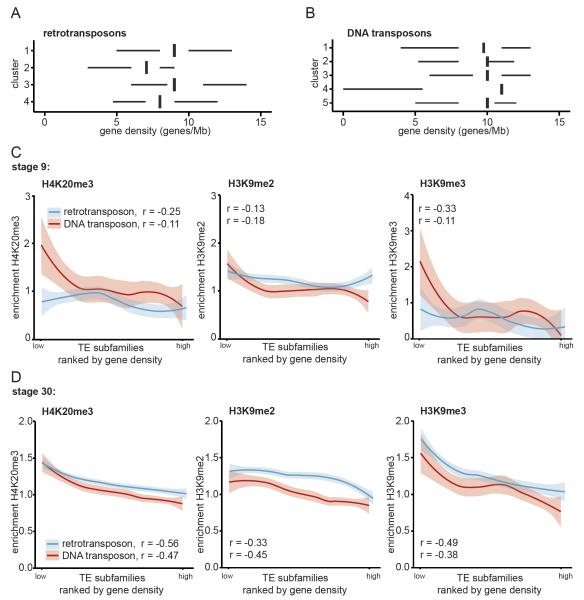
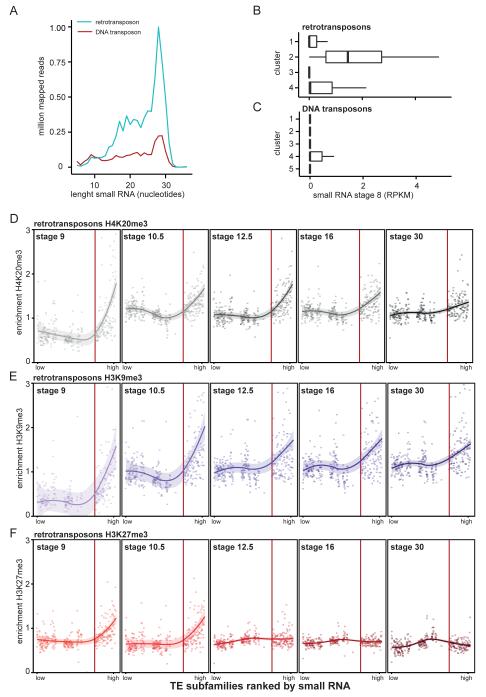
Transposable elements are parasitic genomic elements that can be deleterious for host gene function and genome integrity. Heterochromatic histone modifications are involved in the repression of transposons. However, it remains unknown how these histone modifications mark different types of transposons during embryonic development. Here we document the variety of heterochromatic epigenetic signatures at parasitic elements during development in Xenopus tropicalis, using genome-wide ChIP-sequencing data and ChIP-qPCR analysis. We show that specific subsets of transposons in various families and subfamilies are marked by different combinations of the heterochromatic histone modifications H4K20me3, H3K9me2/3 and H3K27me3. Many DNA transposons are marked at the blastula stage already, whereas at retrotransposons the histone modifications generally accumulate at the gastrula stage or later. Furthermore, transposons marked by H3K9me3 and H4K20me3 are more prominent in gene deserts. Using intra-subfamily divergence as a proxy for age, we show that relatively young DNA transposons are preferentially marked by early embryonic H4K20me3 and H3K27me3. In contrast, relatively young retrotransposons are marked by increasing H3K9me3 and H4K20me3 during development, and are also linked to piRNA-sized small non-coding RNAs. Our results implicate distinct repression mechanisms that operate in a transposon-selective and developmental stage-specific fashion.
转座元件是寄生性基因组元件,可能对宿主基因功能和基因组完整性有害。异染色质组蛋白修饰参与转座子的抑制。然而,在胚胎发育过程中,这些组蛋白修饰如何标记不同类型的转座子仍不清楚。在这里,我们利用全基因组ChIP测序数据和ChIP-qPCR分析,记录了热带爪蟾发育过程中寄生元件处异染色质表观遗传特征的多样性。我们表明,不同家族和亚家族中转座子的特定子集由异染色质组蛋白修饰H4K20me3、H3K9me2/3和H3K27me3的不同组合标记。许多DNA转座子在囊胚期就已被标记,而反转录转座子的组蛋白修饰通常在原肠胚期或更晚的时候积累。此外,由H3K9me3和H4K20me3标记的转座子在基因沙漠中更为突出。利用亚家族内的差异作为年龄的代表,我们表明相对年轻的DNA转座子优先被早期胚胎H4K20me3和H3K27me3标记。相反,相对年轻的反转录转座子在发育过程中被增加的H3K9me3和H4K20me3标记,并且也与piRNA大小的小非编码RNA相关。我们的结果暗示了以转座子选择性和发育阶段特异性方式运作的不同抑制机制。