Levendosky Robert F, Sabantsev Anton, Deindl Sebastian, Bowman Gregory D
T.C. Jenkins Department of Biophysics, Johns Hopkins University, Baltimore, United States.
Department of Cell and Molecular Biology, Science for Life Laboratory, Uppsala University, Uppsala, Sweden.
Elife. 2016 Dec 29;5:e21356. doi: 10.7554/eLife.21356.
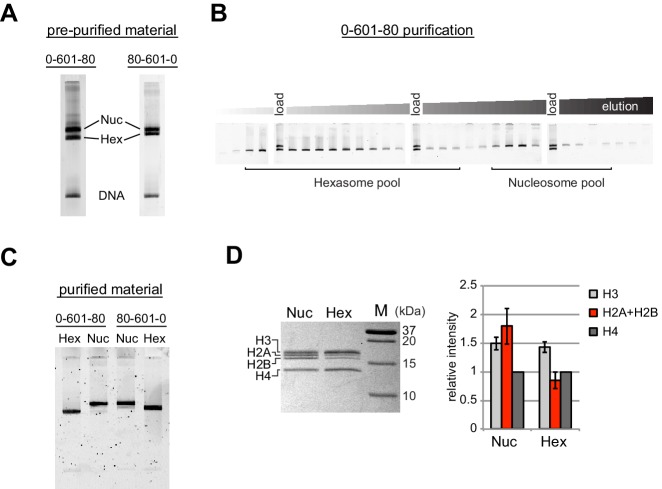
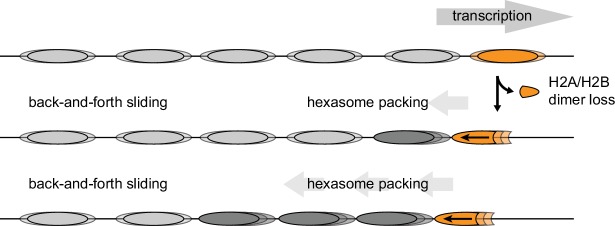
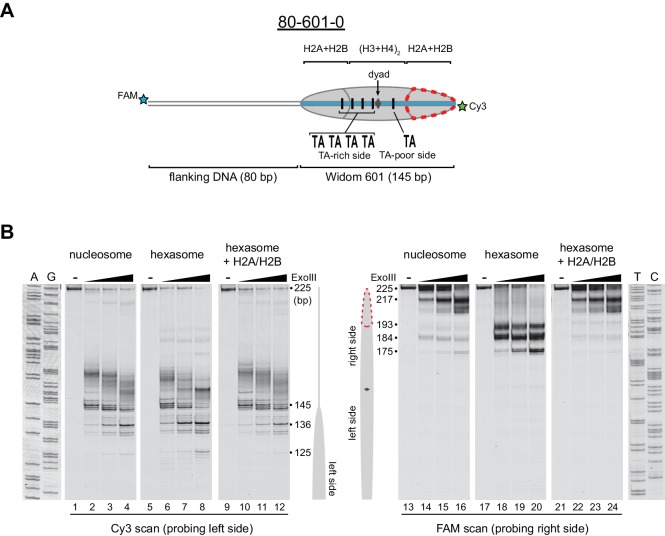
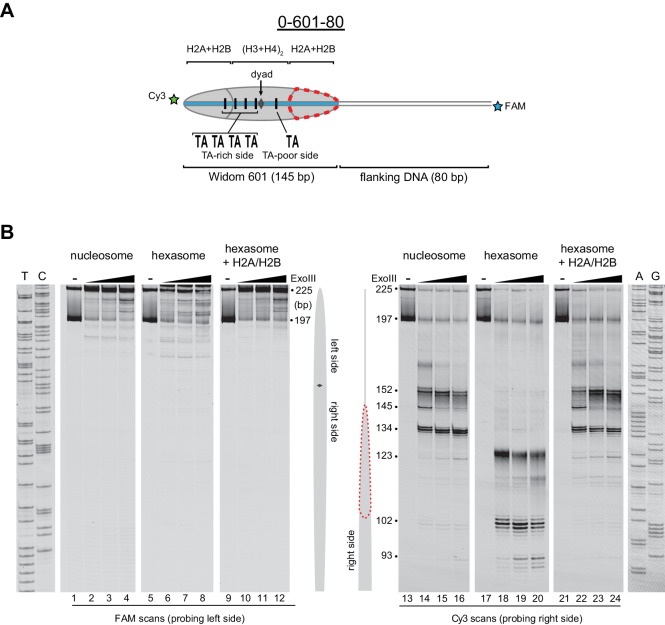
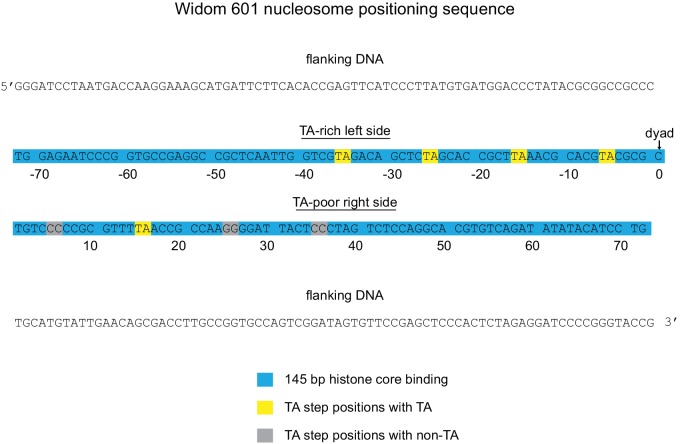
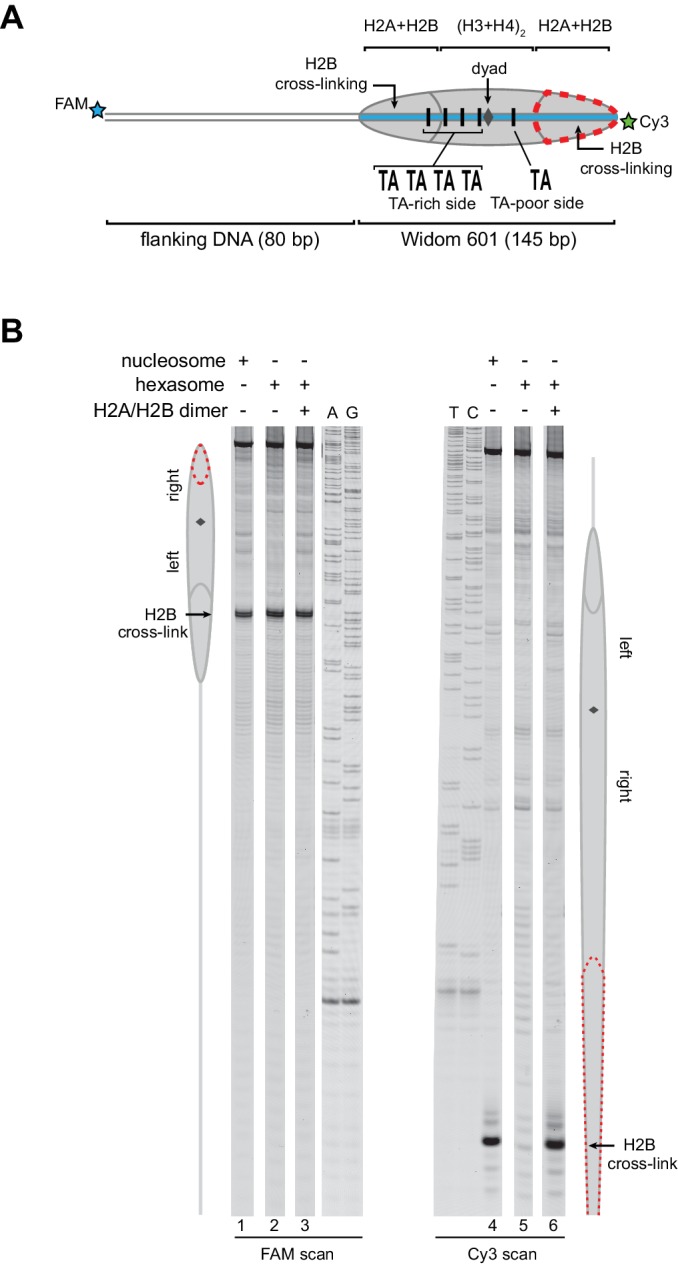
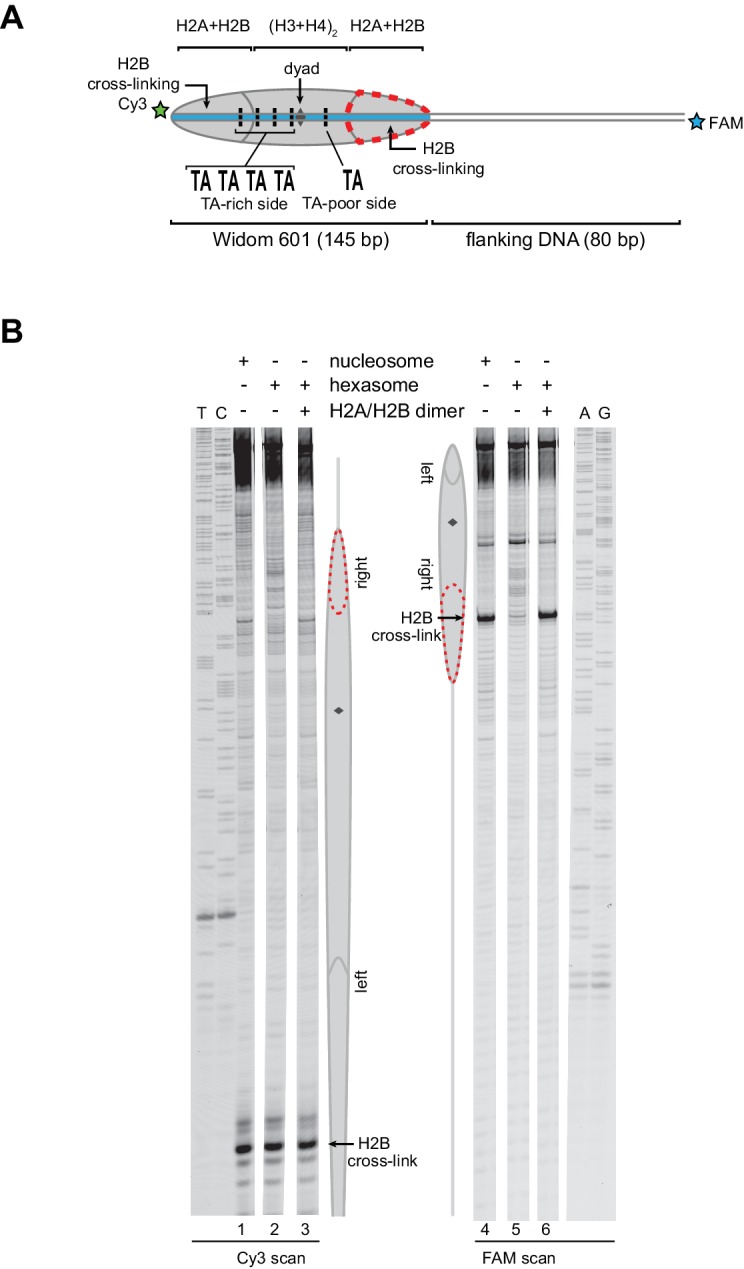
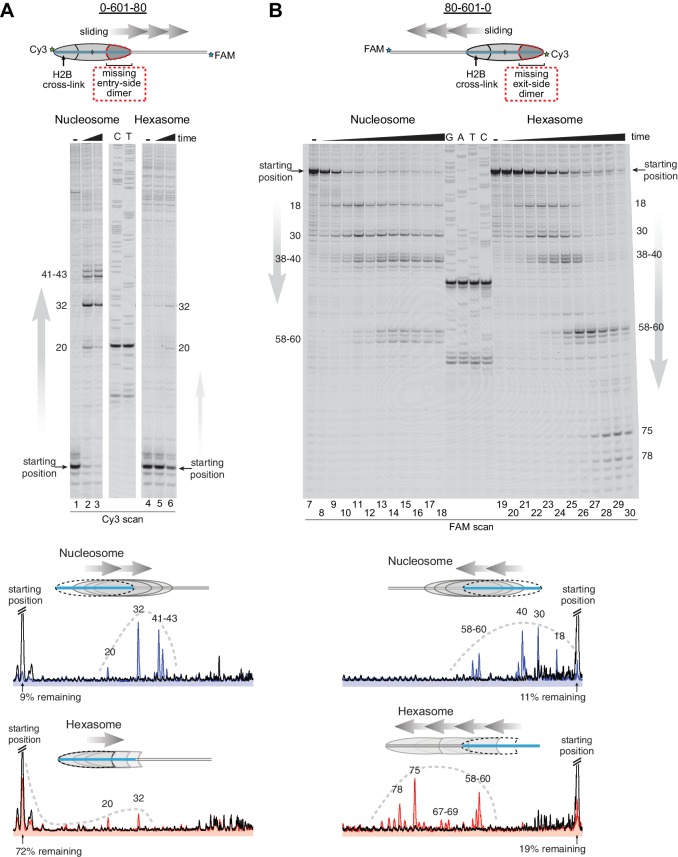
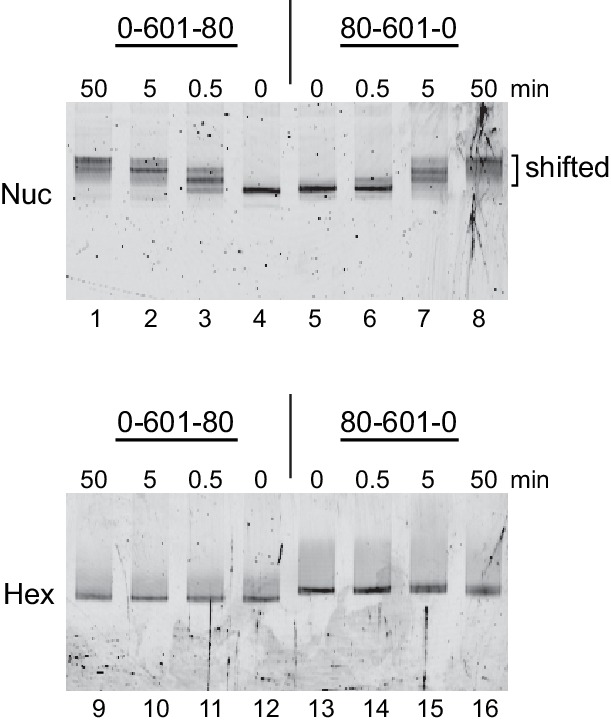
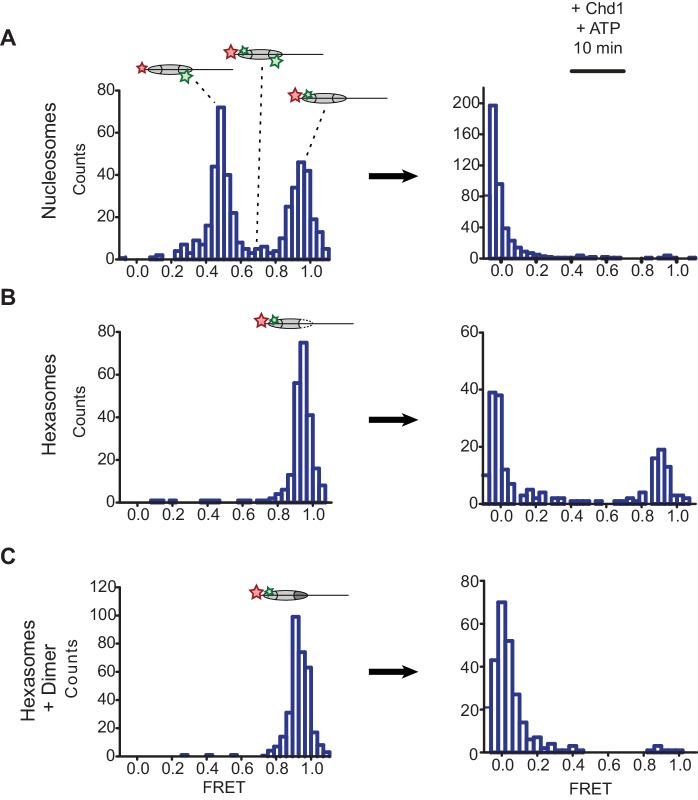
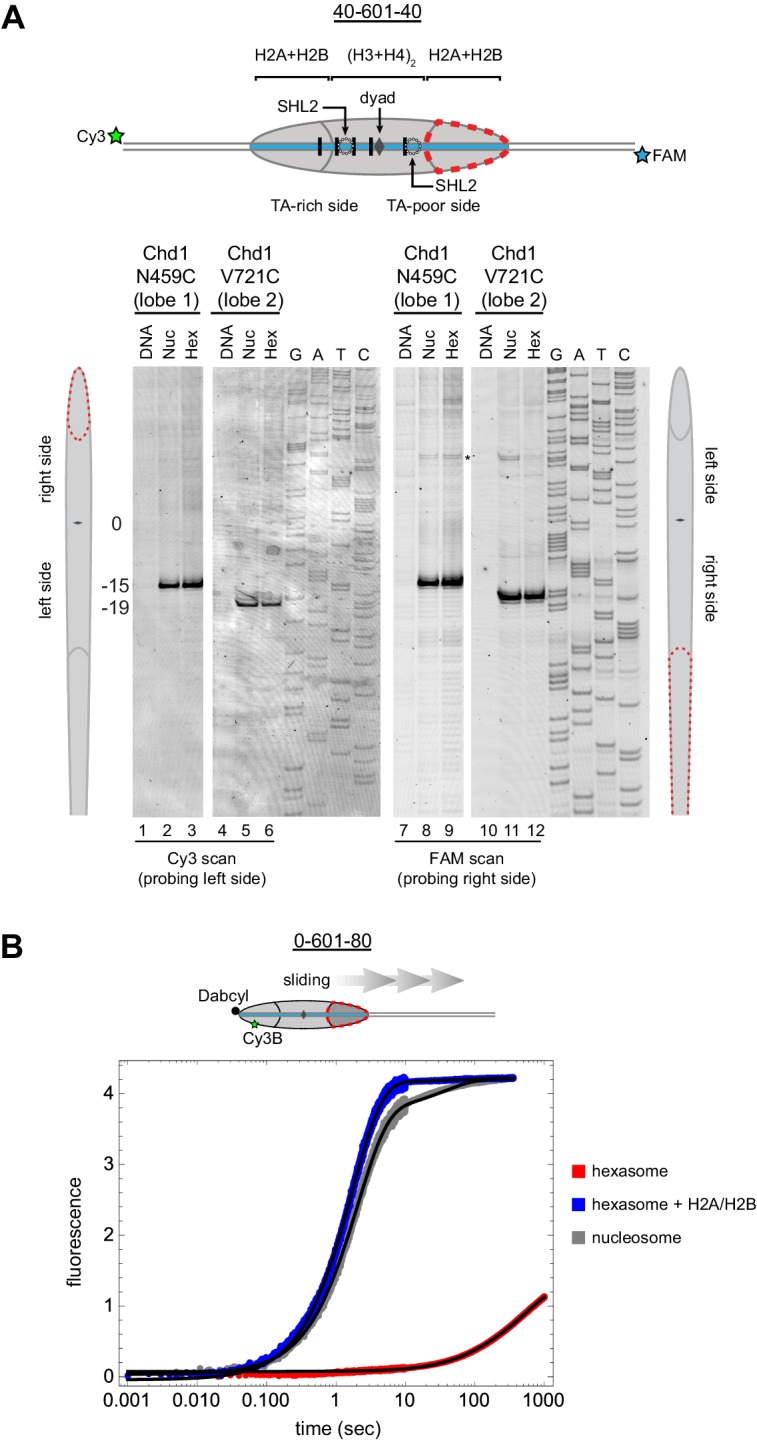
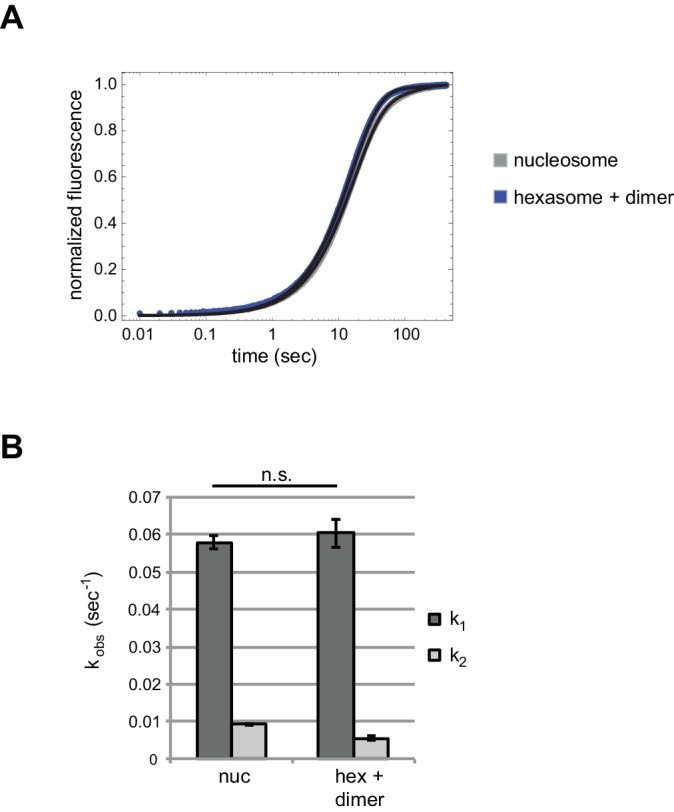
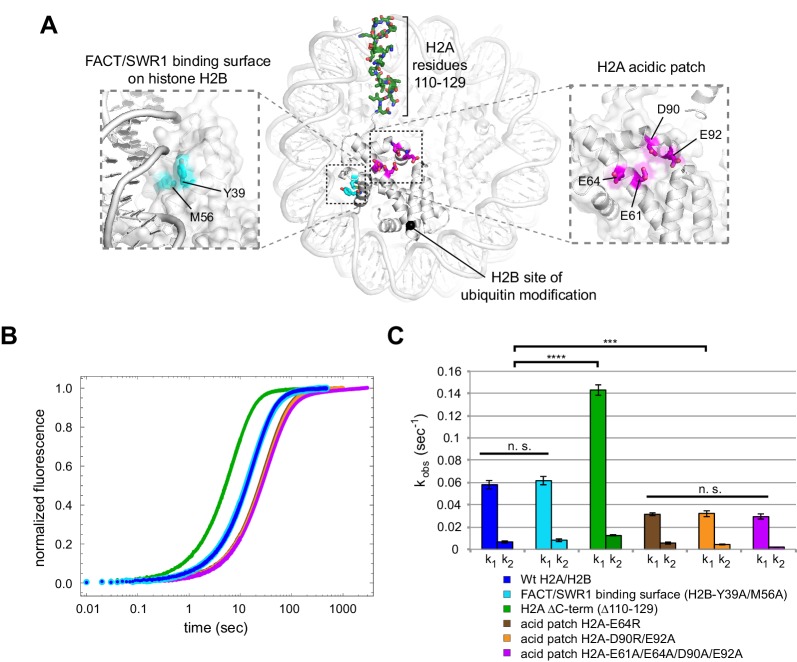
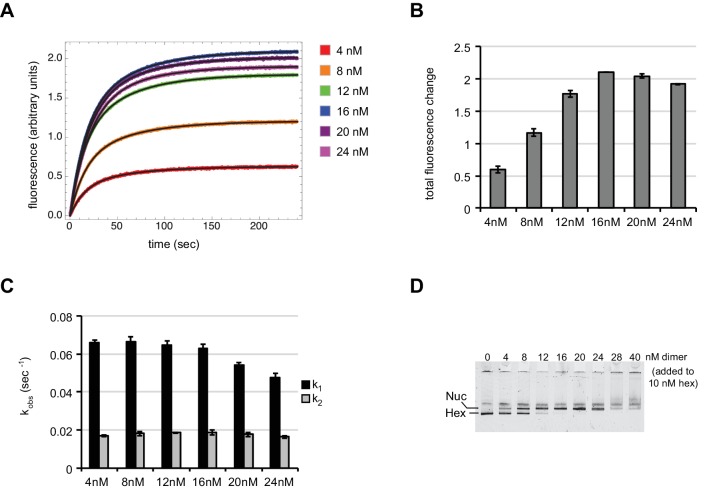
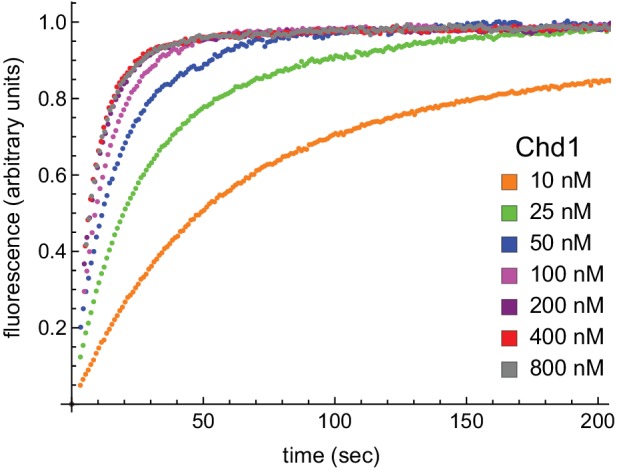
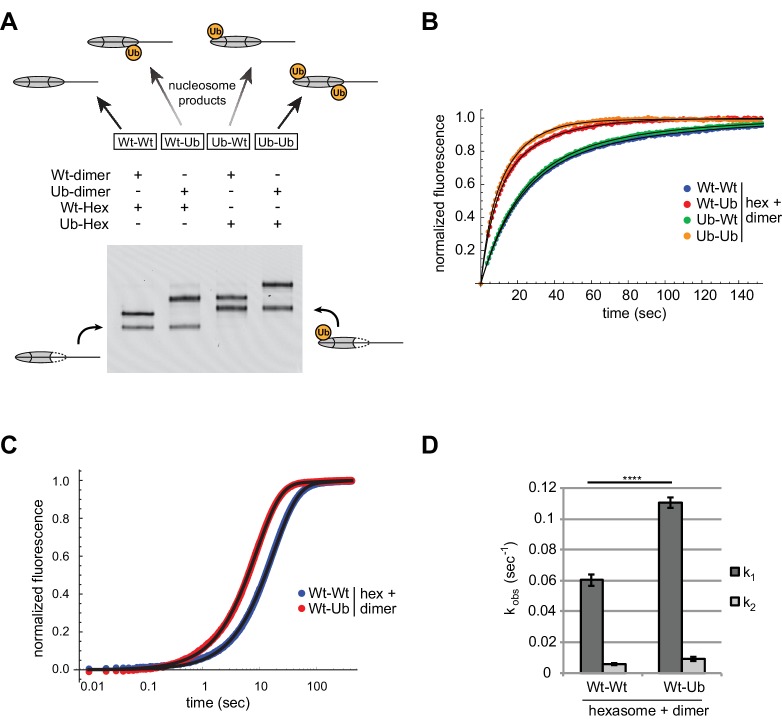
Despite their canonical two-fold symmetry, nucleosomes in biological contexts are often asymmetric: functionalized with post-translational modifications (PTMs), substituted with histone variants, and even lacking H2A/H2B dimers. Here we show that the Widom 601 nucleosome positioning sequence can produce hexasomes in a specific orientation on DNA, providing a useful tool for interrogating chromatin enzymes and allowing for the generation of nucleosomes with precisely defined asymmetry. Using this methodology, we demonstrate that the Chd1 chromatin remodeler from requires H2A/H2B on the entry side for sliding, and thus, unlike the back-and-forth sliding observed for nucleosomes, Chd1 shifts hexasomes unidirectionally. Chd1 takes part in chromatin reorganization surrounding transcribing RNA polymerase II (Pol II), and using asymmetric nucleosomes we show that ubiquitin-conjugated H2B on the entry side stimulates nucleosome sliding by Chd1. We speculate that biased nucleosome and hexasome sliding due to asymmetry contributes to the packing of arrays observed in vivo.
尽管核小体具有典型的二重对称性,但在生物学环境中它们通常是不对称的:通过翻译后修饰(PTM)进行功能化,被组蛋白变体取代,甚至缺乏H2A/H2B二聚体。在这里,我们表明Widom 601核小体定位序列可以在DNA上以特定方向产生六聚体,为研究染色质酶提供了一个有用的工具,并允许生成具有精确界定不对称性的核小体。使用这种方法,我们证明来自……的Chd1染色质重塑因子在进入侧进行滑动时需要H2A/H2B,因此,与核小体观察到的来回滑动不同,Chd1单向移动六聚体。Chd1参与围绕转录RNA聚合酶II(Pol II)的染色质重组,并且使用不对称核小体我们表明进入侧泛素化的H2B刺激Chd1介导的核小体滑动。我们推测由于不对称导致的偏向性核小体和六聚体滑动有助于体内观察到的阵列堆积。