Coda Davide M, Gaarenstroom Tessa, East Philip, Patel Harshil, Miller Daniel S J, Lobley Anna, Matthews Nik, Stewart Aengus, Hill Caroline S
Developmental Signalling Laboratory, The Francis Crick Institute, London, United Kingdom.
Bioinformatics and Biostatistics, The Francis Crick Institute, London, United Kingdom.
Elife. 2017 Feb 13;6:e22474. doi: 10.7554/eLife.22474.
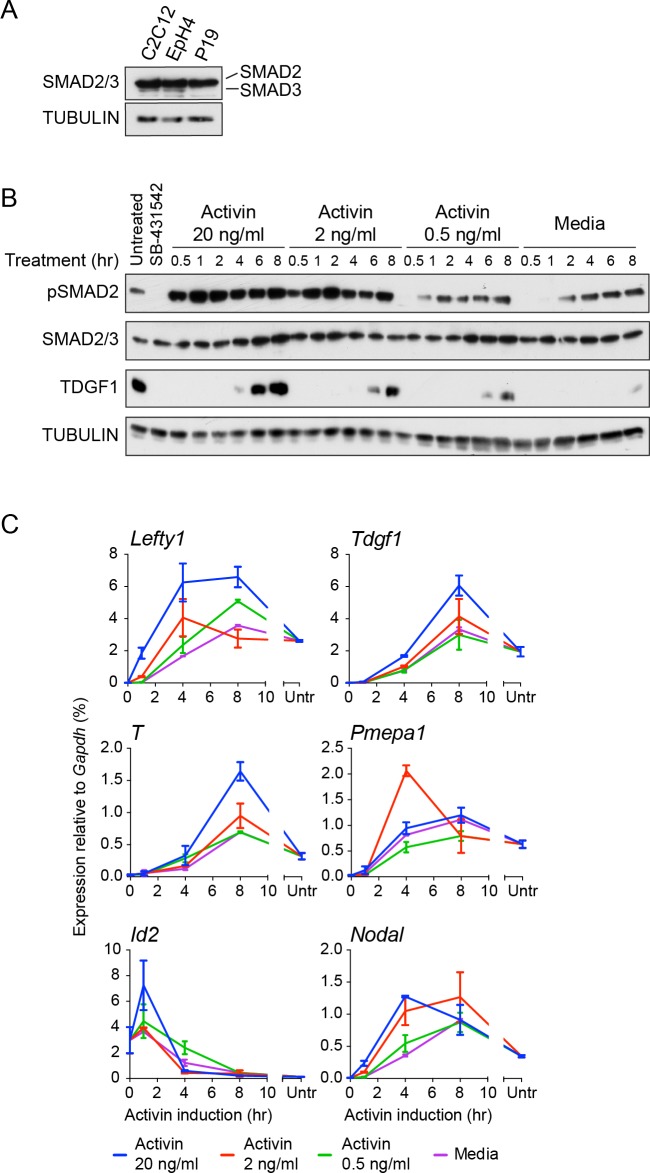
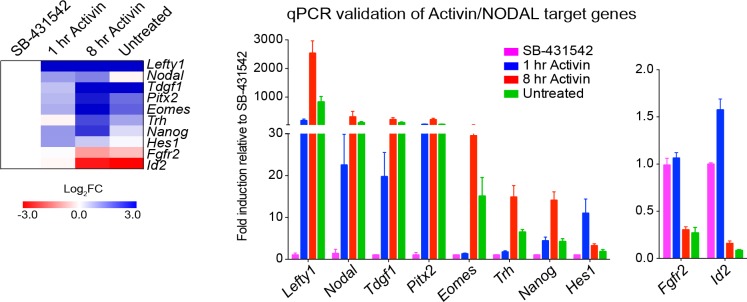
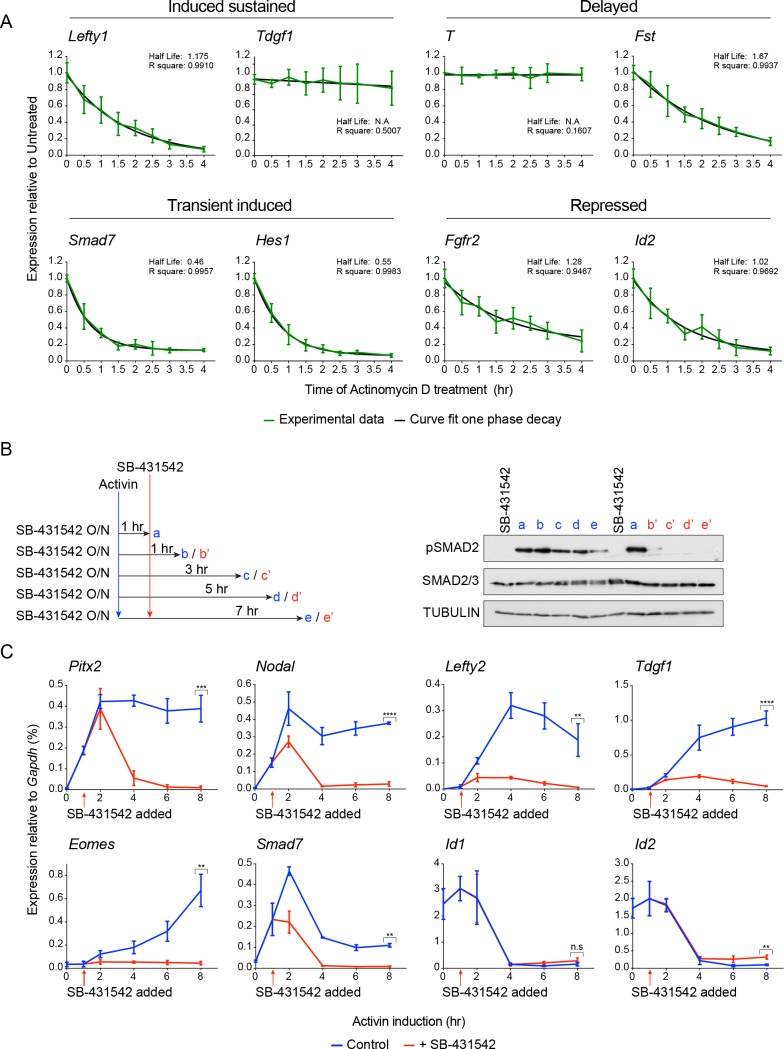
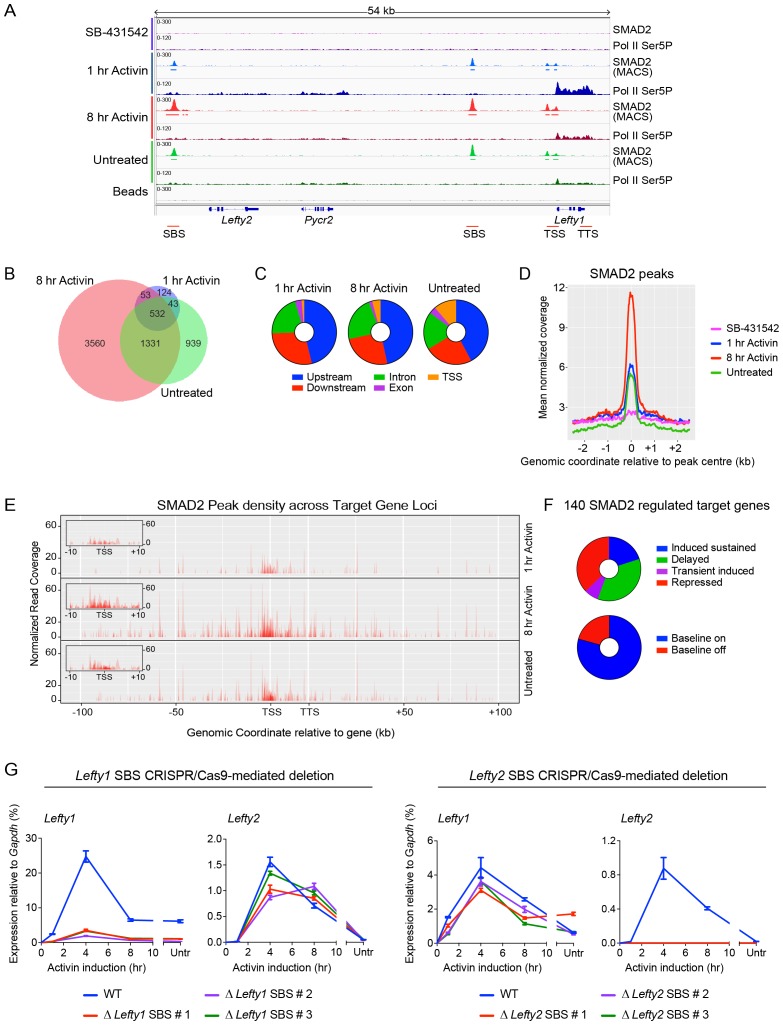
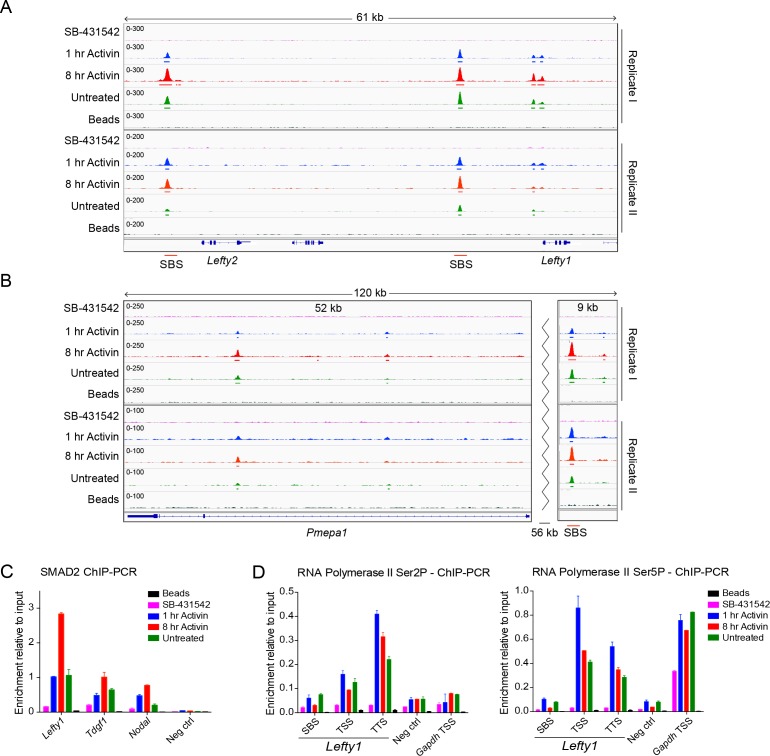
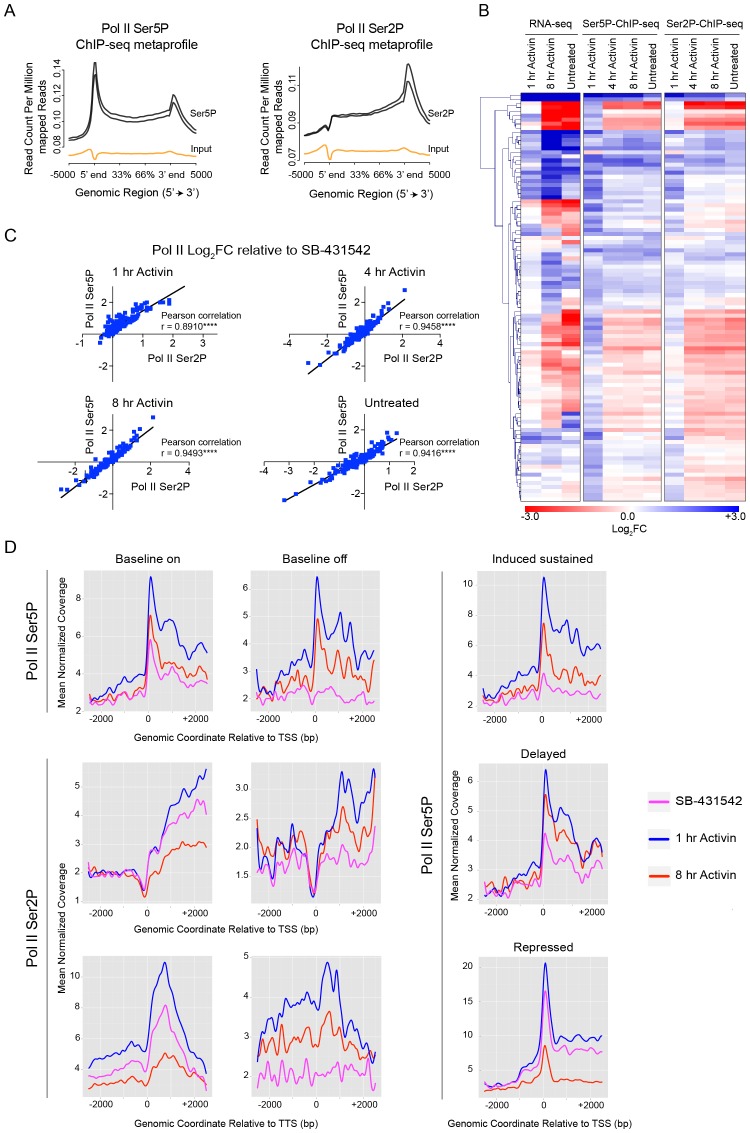
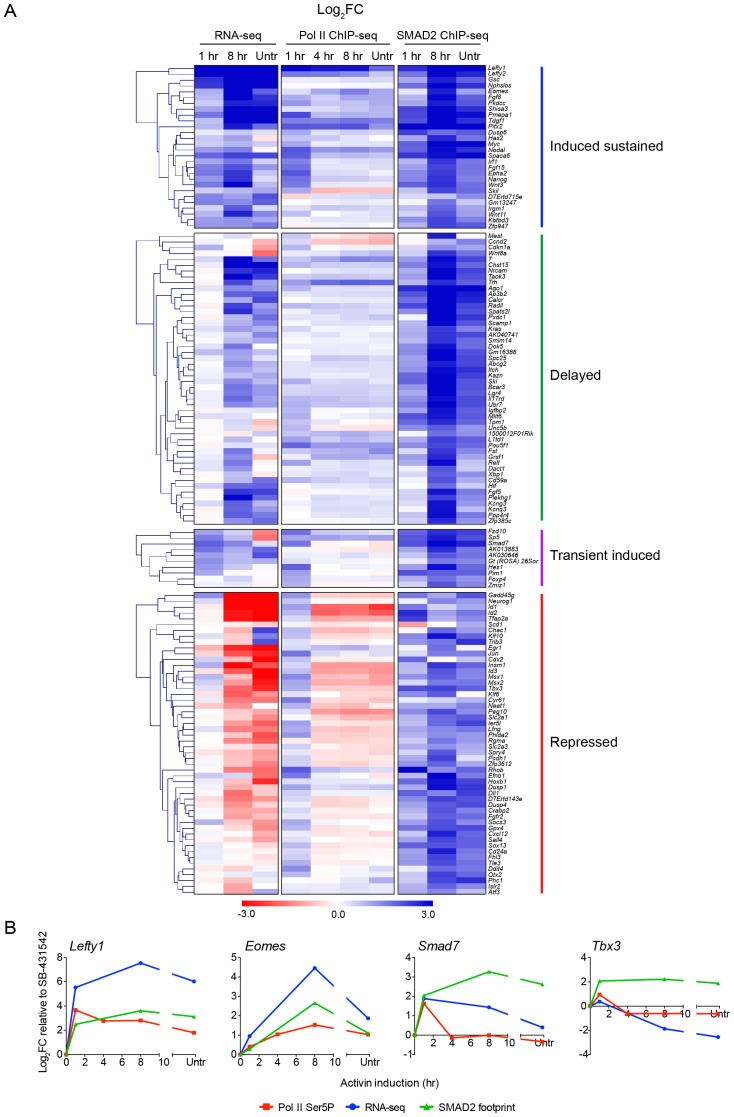
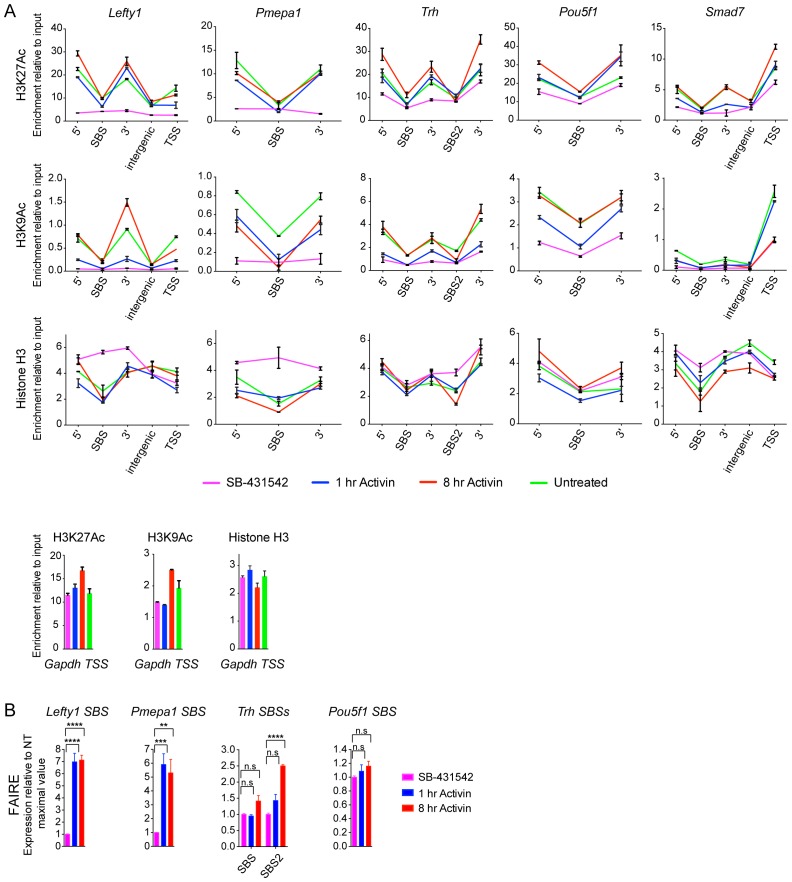
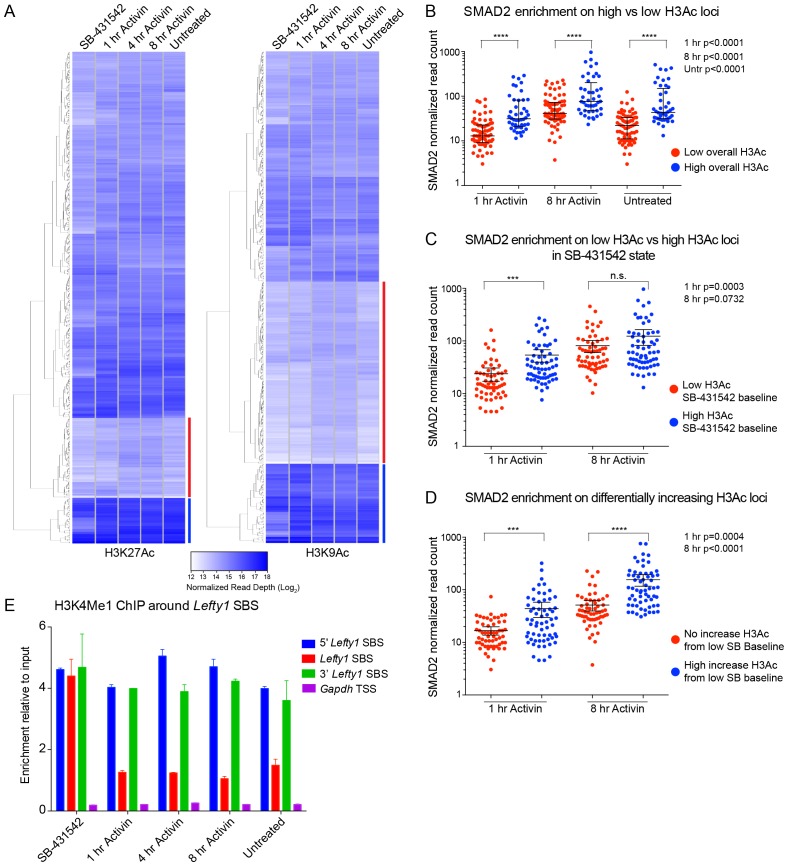
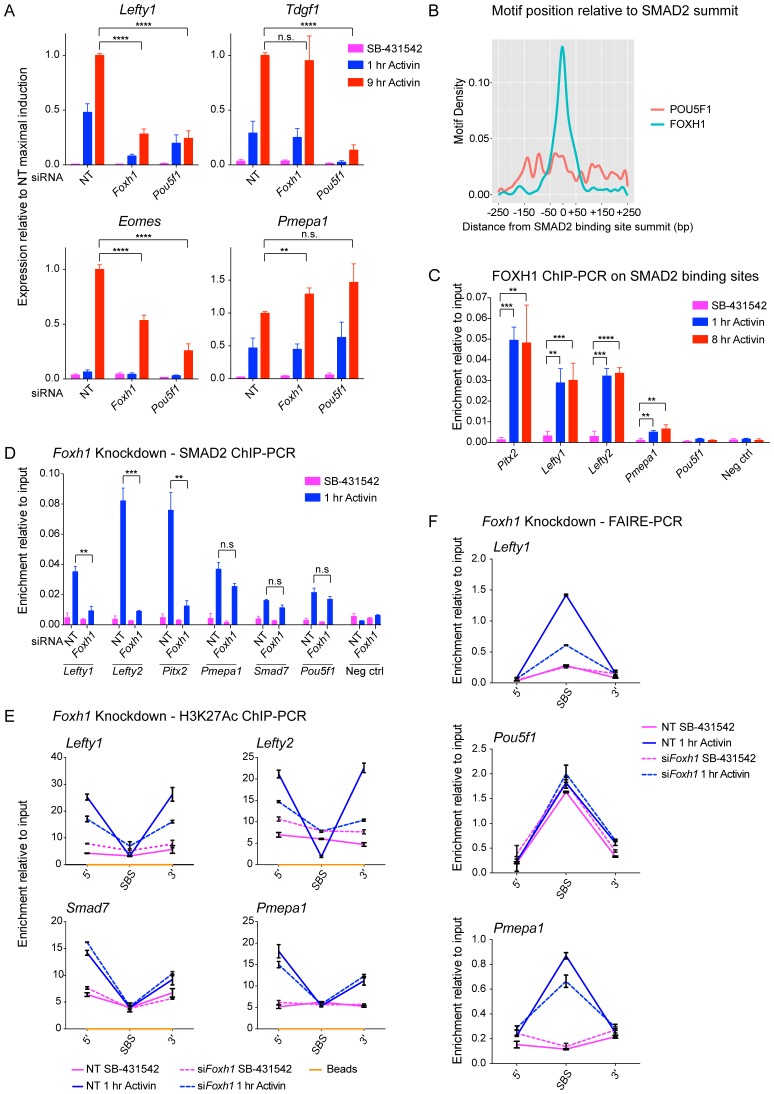
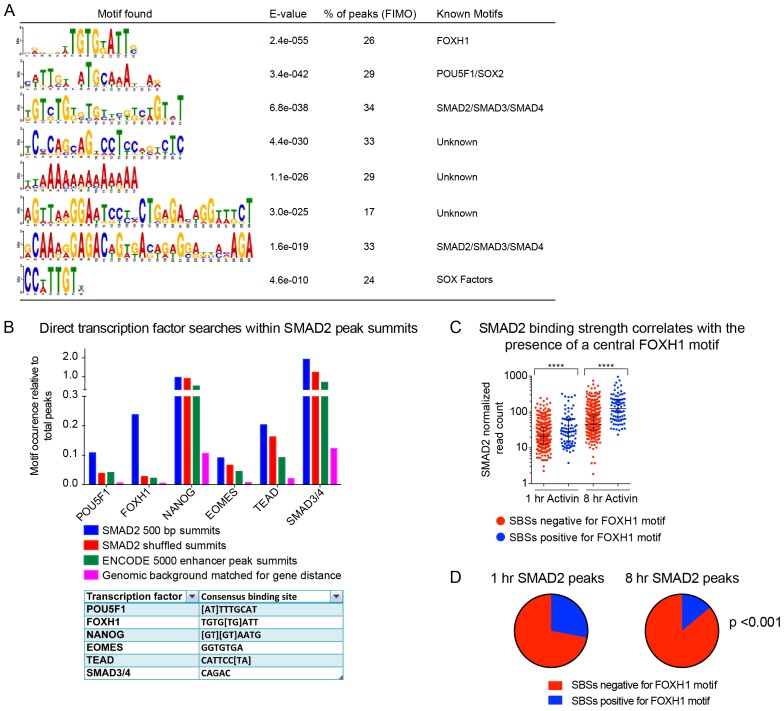
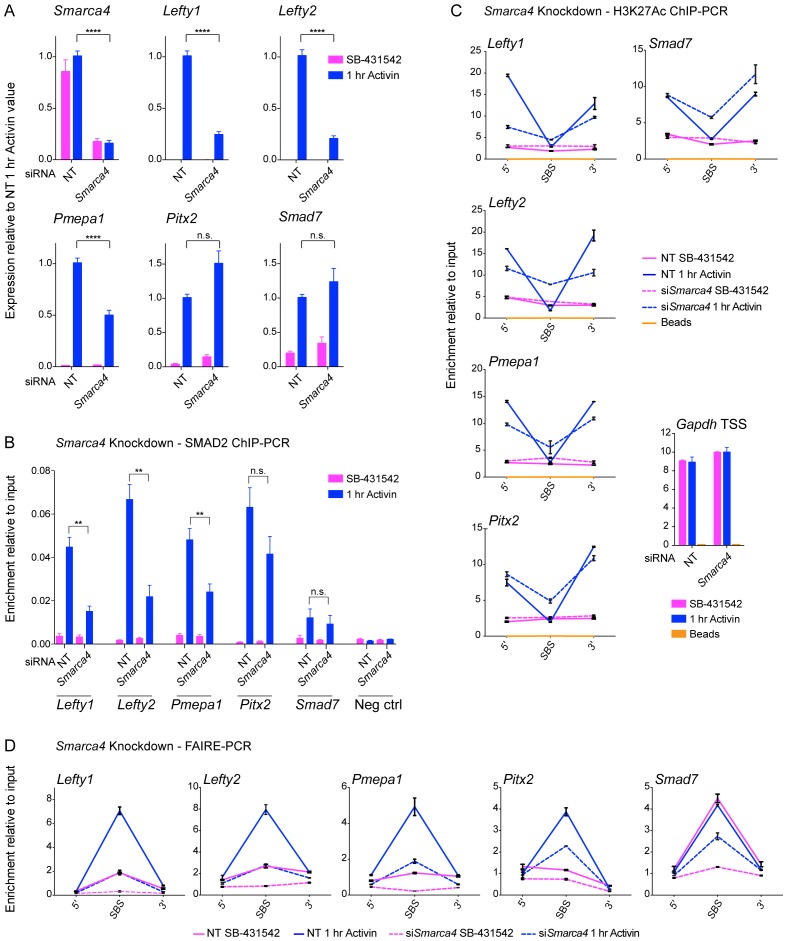
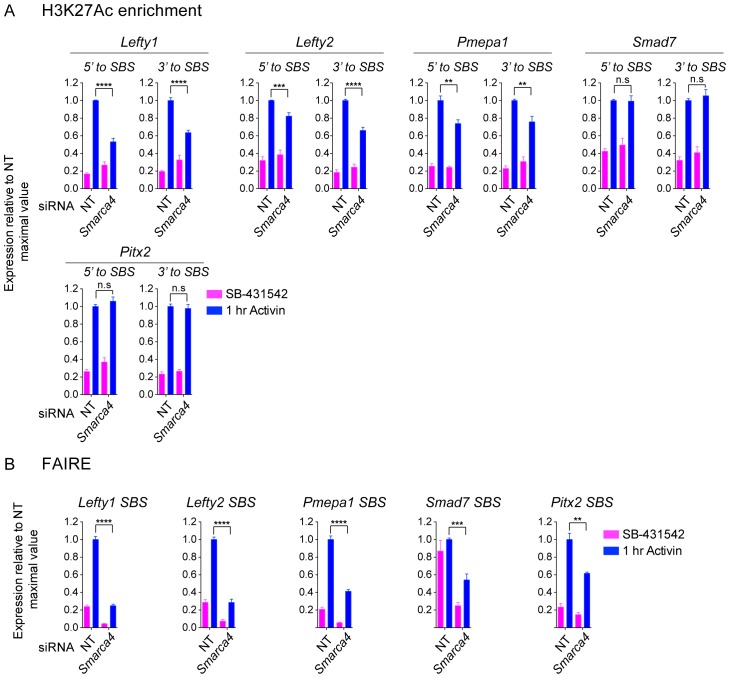
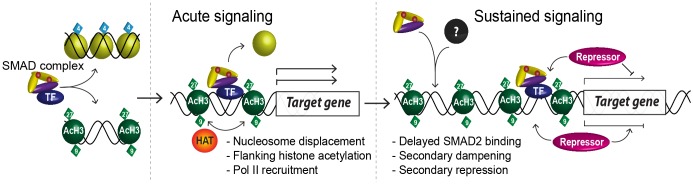
NODAL/Activin signaling orchestrates key processes during embryonic development via SMAD2. How SMAD2 activates programs of gene expression that are modulated over time however, is not known. Here we delineate the sequence of events that occur from SMAD2 binding to transcriptional activation, and the mechanisms underlying them. NODAL/Activin signaling induces dramatic chromatin landscape changes, and a dynamic transcriptional network regulated by SMAD2, acting via multiple mechanisms. Crucially we have discovered two modes of SMAD2 binding. SMAD2 can bind pre-acetylated nucleosome-depleted sites. However, it also binds to unacetylated, closed chromatin, independently of pioneer factors, where it induces nucleosome displacement and histone acetylation. For a subset of genes, this requires SMARCA4. We find that long term modulation of the transcriptional responses requires continued NODAL/Activin signaling. Thus SMAD2 binding does not linearly equate with transcriptional kinetics, and our data suggest that SMAD2 recruits multiple co-factors during sustained signaling to shape the downstream transcriptional program.
NODAL/激活素信号通过SMAD2在胚胎发育过程中协调关键过程。然而,SMAD2如何激活随时间变化而被调控的基因表达程序尚不清楚。在这里,我们描绘了从SMAD2结合到转录激活所发生的一系列事件及其潜在机制。NODAL/激活素信号诱导了显著的染色质景观变化,以及一个由SMAD2通过多种机制调控的动态转录网络。至关重要的是,我们发现了SMAD2结合的两种模式。SMAD2可以结合预乙酰化的核小体缺失位点。然而,它也能独立于先驱因子与未乙酰化的封闭染色质结合,在那里它诱导核小体位移和组蛋白乙酰化。对于一部分基因,这需要SMARCA4。我们发现转录反应的长期调控需要持续的NODAL/激活素信号。因此,SMAD2结合并不与转录动力学呈线性等同,我们的数据表明,SMAD2在持续信号传导过程中招募多种辅助因子来塑造下游转录程序。