Blotnick Edna, Sol Asaf, Bachrach Gilad, Muhlrad Andras
Department of Medical Neurobiology, Institute for Medical Research-Israel-Canada, Hebrew University of Jerusalem, Jerusalem, Israel.
Institute of Dental Sciences, Hebrew University-Hadassah School of Dental Medicine, Jerusalem, Israel.
BMC Biochem. 2017 Mar 6;18(1):3. doi: 10.1186/s12858-017-0078-0.
Histatins are histidine rich polypeptides produced in the parotid and submandibular gland and secreted into the saliva. Histatin-3 and -5 are the most important polycationic histatins. They possess antimicrobial activity against fungi such as Candida albicans. Histatin-5 has a higher antifungal activity than histatin-3 while histatin-3 is mostly involved in wound healing in the oral cavity. We found that these histatins, like other polycationic peptides and proteins, such as LL-37, lysozyme and histones, interact with extracellular actin.
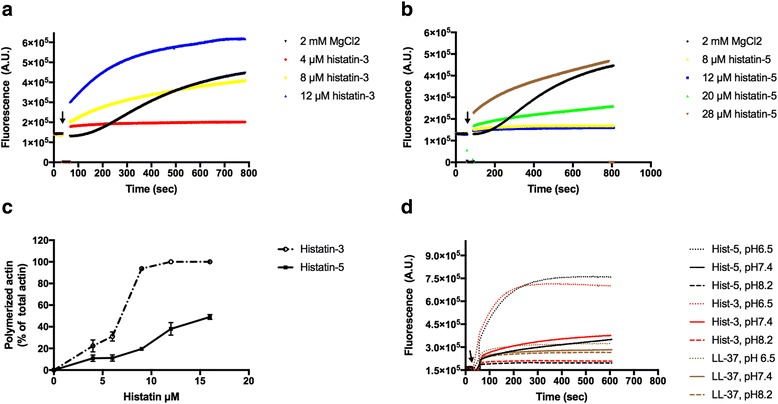
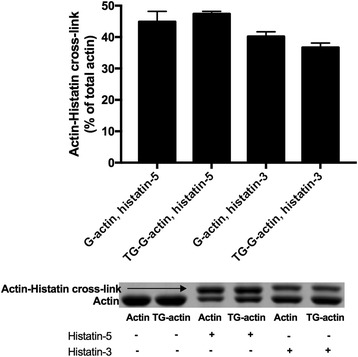
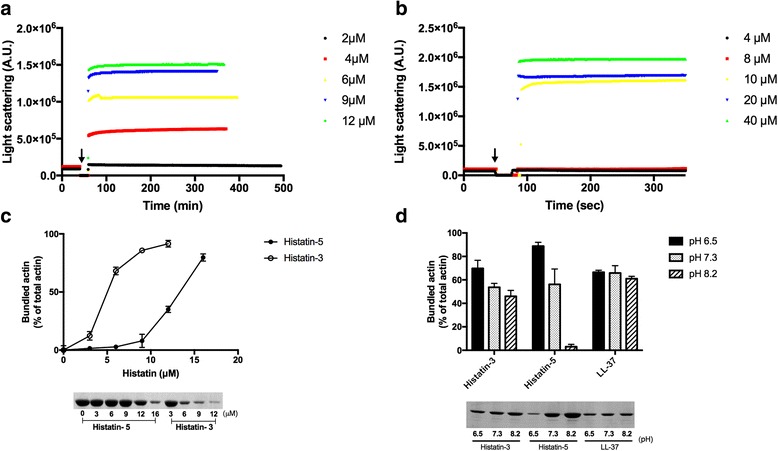
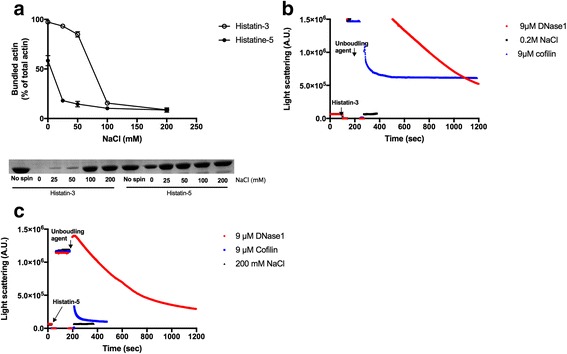
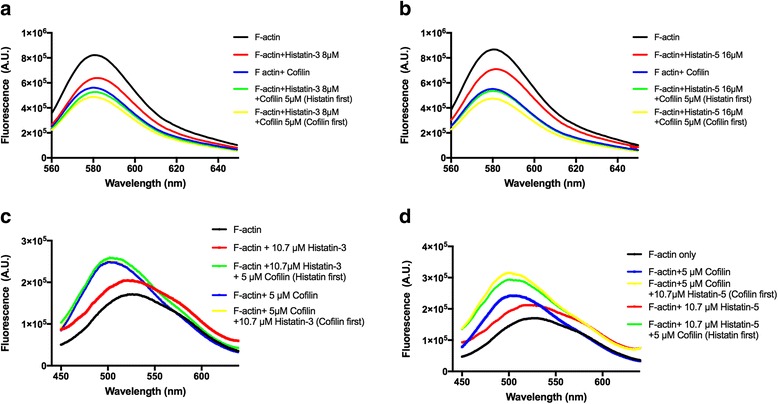
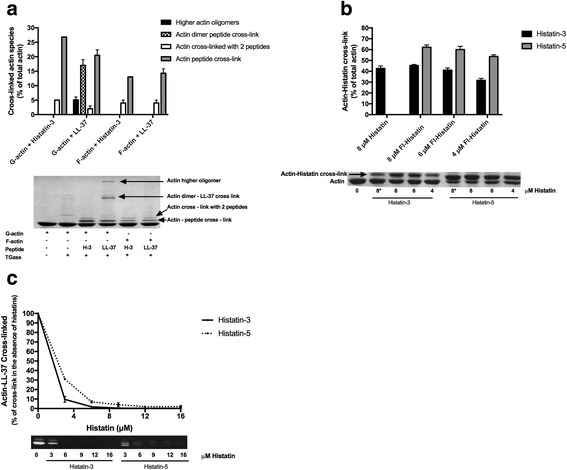
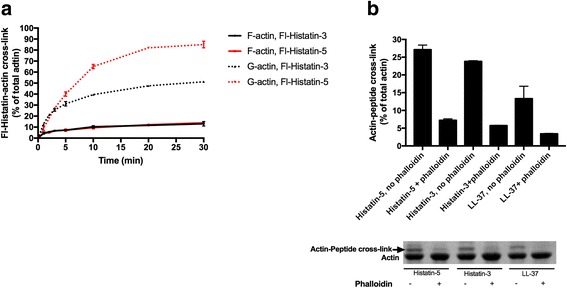
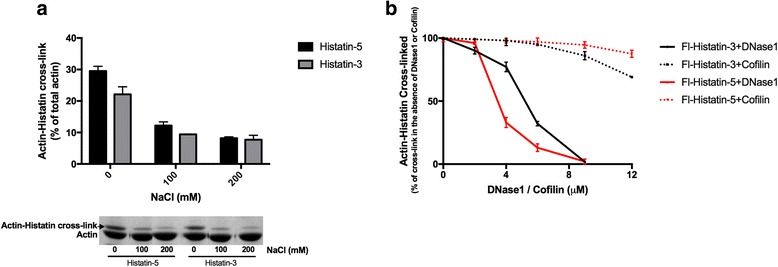
Histatin-3 and -5 polymerize globular actin (G-actin) to filamentous actin (F-actin) and bundle F-actin filaments. Both actin polymerization and bundling by histatins is pH sensitive due to the high histidine content of histatins. In spite of the equal number of net positive charges and histidine residues in histatin-3 and -5, less histatin-3 is needed than histatin-5 for polymerization and bundling of actin. The efficiency of actin polymerization and bundling by histatins greatly increases with decreasing pH. Histatin-3 and -5 induced actin bundles are dissociated by 100 and 50 mM NaCl, respectively. The relatively low NaCl concentration required to dissociate histatin-induced bundles implies that the actin-histatin filaments bind to each other mainly by electrostatic forces. The binding of histatin-3 to F-actin is stronger than that of histatin-5 showing that hydrophobic forces have also some role in histatin-3- actin interaction. Histatins affect the fluorescence of probes attached to the D-loop of G-actin indicating histatin induced changes in actin structure. Transglutaminase cross-links histatins to actin. Competition and limited proteolysis experiments indicate that the main histatin cross-linking site on actin is glutamine-49 on the D-loop of actin.
Both histatin-3 and -5 interacts with actin, however, histatin 3 binds stronger to actin and affects actin structure at lower concentration than histatin-5 due to the extra 8 amino acid sequence at the C-terminus of histatin-3. Extracellular actin might regulate histatin activity in the oral cavity, which should be the subject of further investigation.
富组蛋白是在腮腺和颌下腺产生并分泌到唾液中的富含组氨酸的多肽。组蛋白-3和-5是最重要的聚阳离子富组蛋白。它们对白色念珠菌等真菌具有抗菌活性。组蛋白-5的抗真菌活性高于组蛋白-3,而组蛋白-3主要参与口腔伤口愈合。我们发现,这些富组蛋白与其他聚阳离子肽和蛋白质,如LL-37、溶菌酶和组蛋白一样,与细胞外肌动蛋白相互作用。
组蛋白-3和-5将球状肌动蛋白(G-肌动蛋白)聚合成丝状肌动蛋白(F-肌动蛋白)并使F-肌动蛋白丝成束。由于富组蛋白的组氨酸含量高,富组蛋白介导的肌动蛋白聚合和成束均对pH敏感。尽管组蛋白-3和-5的净正电荷数和组氨酸残基数量相等,但肌动蛋白聚合和成束所需的组蛋白-3比组蛋白-5少。随着pH降低,富组蛋白介导的肌动蛋白聚合和成束效率大大提高。组蛋白-3和-5诱导的肌动蛋白束分别在100和50 mM NaCl作用下解聚。解离富组蛋白诱导的肌动蛋白束所需的相对较低NaCl浓度表明,肌动蛋白-富组蛋白丝主要通过静电力相互结合。组蛋白-3与F-肌动蛋白的结合比组蛋白-5更强,表明疏水作用力在组蛋白-3与肌动蛋白的相互作用中也起一定作用。富组蛋白影响附着在G-肌动蛋白D环上的探针的荧光,表明富组蛋白诱导了肌动蛋白结构的变化。转谷氨酰胺酶使富组蛋白与肌动蛋白交联。竞争和有限蛋白酶解实验表明,肌动蛋白上主要的富组蛋白交联位点是肌动蛋白D环上的谷氨酰胺-49。
组蛋白-3和-5均与肌动蛋白相互作用,然而,由于组蛋白-3的C末端有额外的8个氨基酸序列,组蛋白3与肌动蛋白的结合更强,且在比组蛋白-5更低的浓度下就能影响肌动蛋白结构。细胞外肌动蛋白可能调节口腔中富组蛋白的活性,这应是进一步研究的课题。