Aguilera Elizabeth R, Erickson Andrea K, Jesudhasan Palmy R, Robinson Christopher M, Pfeiffer Julie K
Department of Microbiology, University of Texas Southwestern Medical Center, Dallas, Texas, USA.
Department of Microbiology, University of Texas Southwestern Medical Center, Dallas, Texas, USA
mBio. 2017 Mar 14;8(2):e02020-16. doi: 10.1128/mBio.02020-16.
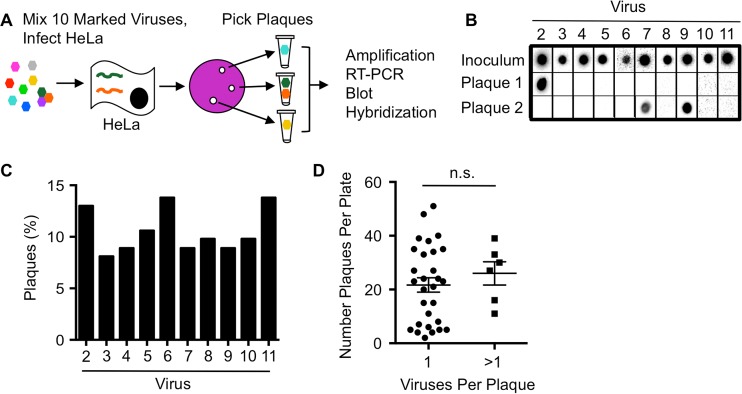
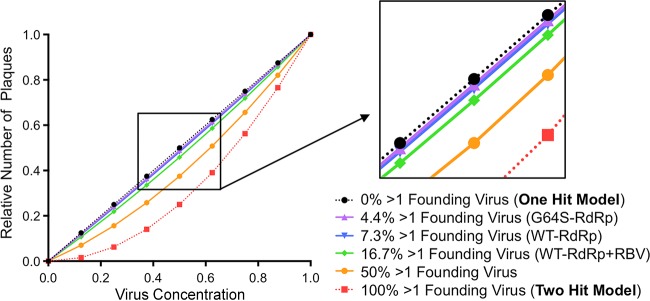
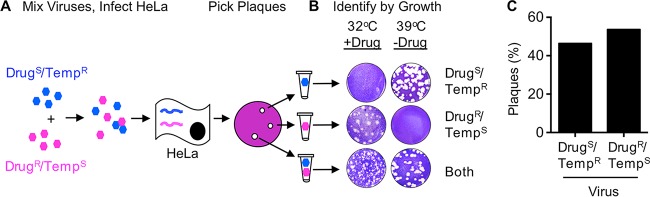
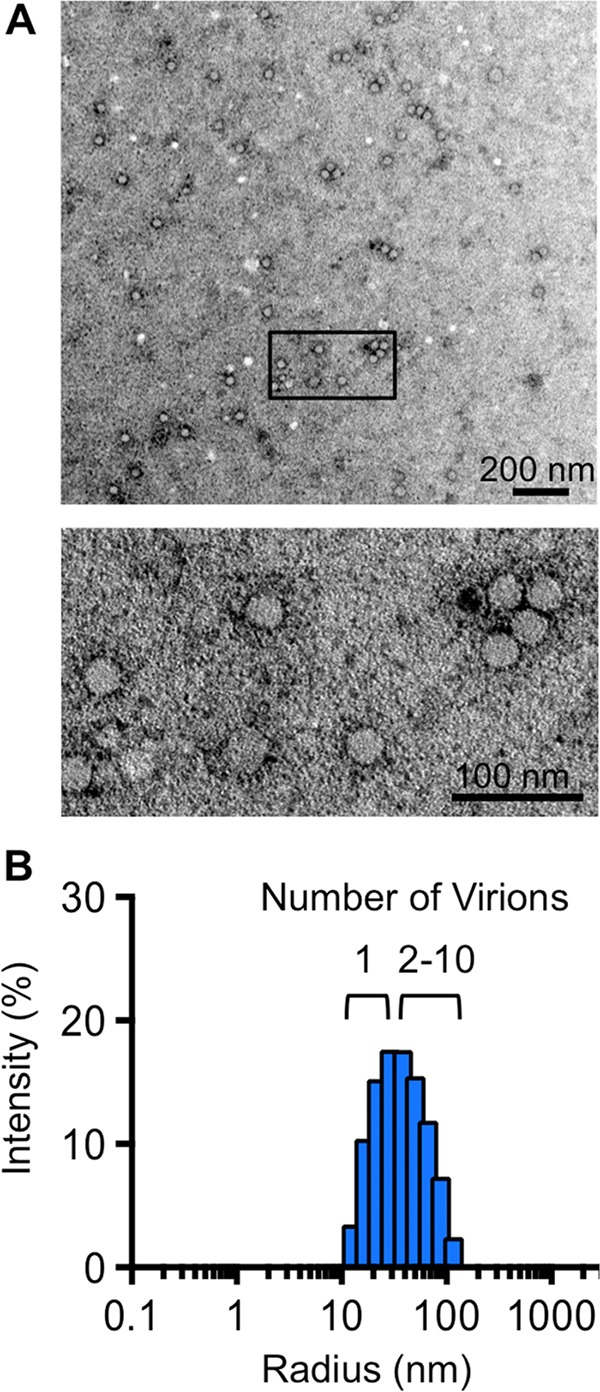
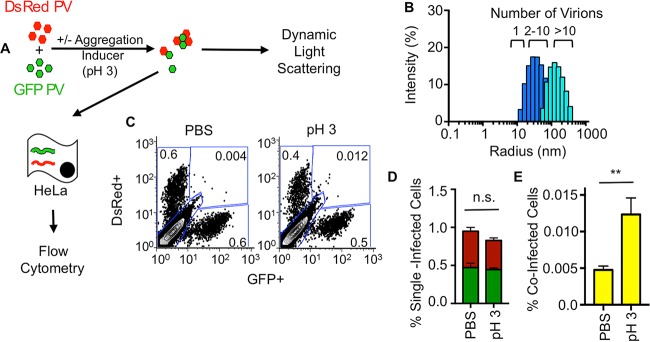
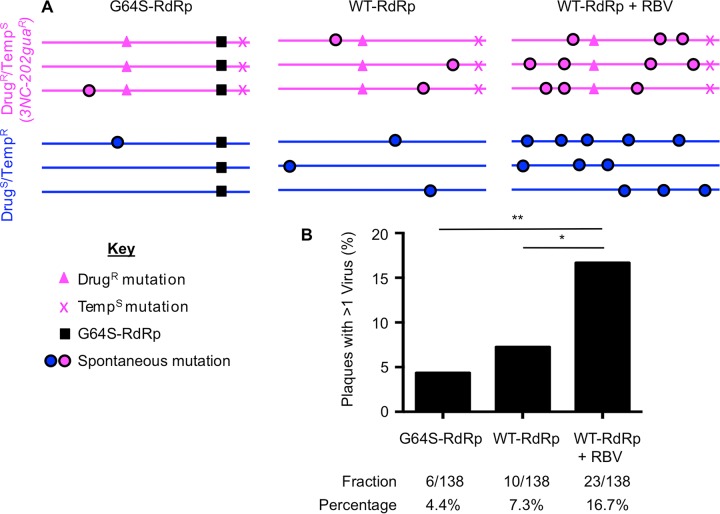
The plaque assay is a common technique used to measure virus concentrations and is based upon the principle that each plaque represents a single infectious unit. As such, the number of plaques is expected to correlate linearly with the virus dilution plated, and each plaque should be formed by a single founder virus. Here, we examined whether more than one virus can contribute to plaque formation. By using genetic and phenotypic assays with genetically marked polioviruses, we found that multiple parental viruses are present in 5 to 7% of plaques, even at an extremely low multiplicity of infection. We demonstrated through visual and biophysical assays that, like many viral stocks, our viral stocks contain both single particles and aggregates. These data suggest that aggregated virions are capable of inducing coinfection and chimeric plaque formation. In fact, inducing virion aggregation via exposure to low pH increased coinfection in a flow cytometry-based assay. We hypothesized that plaques generated by viruses with high mutation loads may have higher coinfection frequencies due to processes restoring fitness, such as complementation and recombination. Indeed, we found that coinfection frequency correlated with mutation load, with 17% chimeric plaque formation for heavily mutagenized viruses. Importantly, the frequency of chimeric plaques may be underestimated by up to threefold, since coinfection with the same parental virus cannot be scored in our assay. This work indicates that more than one virus can contribute to plaque formation and that coinfection may assist plaque formation in situations where the amount of genome damage is high. One of the most common methods to quantify viruses is the plaque assay, where it is generally presumed that each plaque represents a single infectious virus. Using genetically marked polioviruses, we demonstrate that a plaque can contain more than one parental virus, likely due to aggregates within virus stocks that induce coinfection of a cell. A relatively small number of plaques are the products of coinfection for our standard virus stocks. However, mutagenized virus stocks with increased genome damage give rise to a higher amount of plaques that are chimeric. These results suggest that coinfection may aid plaque formation of viruses with genome damage, possibly due to processes such as complementation and recombination. Overall, our results suggest that the relationship between viral dilution and plaque number may not be linear, particularly for mutagenized viral populations.
蚀斑测定法是一种用于测量病毒浓度的常用技术,其基于每个蚀斑代表一个单一感染单位的原理。因此,蚀斑数量预计与接种的病毒稀释度呈线性相关,并且每个蚀斑应由单个起始病毒形成。在这里,我们研究了是否有不止一种病毒能促成蚀斑形成。通过对带有遗传标记的脊髓灰质炎病毒进行遗传和表型分析,我们发现即使在极低的感染复数下,5%至7%的蚀斑中存在多种亲代病毒。我们通过视觉和生物物理分析证明,与许多病毒原液一样,我们的病毒原液既包含单个病毒粒子,也包含聚集体。这些数据表明,聚集的病毒粒子能够诱导共感染和嵌合蚀斑形成。事实上,在基于流式细胞术的分析中,通过暴露于低pH值诱导病毒粒子聚集会增加共感染。我们推测,由于恢复适应性的过程,如互补和重组,具有高突变负荷的病毒产生的蚀斑可能具有更高的共感染频率。确实,我们发现共感染频率与突变负荷相关,对于高度诱变的病毒,嵌合蚀斑形成率为17%。重要的是,嵌合蚀斑的频率可能被低估多达三倍,因为在我们的分析中无法对与相同亲代病毒的共感染进行计分。这项工作表明不止一种病毒能促成蚀斑形成,并且在基因组损伤量高的情况下,共感染可能有助于蚀斑形成。定量病毒最常用的方法之一是蚀斑测定法,通常假定每个蚀斑代表一个单一感染性病毒。使用带有遗传标记的脊髓灰质炎病毒,我们证明一个蚀斑可以包含不止一种亲代病毒,这可能是由于病毒原液中的聚集体诱导细胞共感染。对于我们的标准病毒原液,相对少量的蚀斑是共感染的产物。然而,基因组损伤增加的诱变病毒原液会产生更高数量的嵌合蚀斑。这些结果表明,共感染可能有助于基因组损伤病毒的蚀斑形成,这可能是由于互补和重组等过程。总体而言,我们的结果表明病毒稀释度与蚀斑数量之间的关系可能不是线性的,特别是对于诱变的病毒群体。