Lin Chuwen, Yao Erica, Zhang Kuan, Jiang Xuan, Croll Stacey, Thompson-Peer Katherine, Chuang Pao-Tien
Cardiovascular Research Institute, University of California, San Francisco, San Francisco, United States.
Department of Physiology, Howard Hughes Medical institute, University of California, San Francisco, San Francisco, United States.
Elife. 2017 Mar 21;6:e21130. doi: 10.7554/eLife.21130.
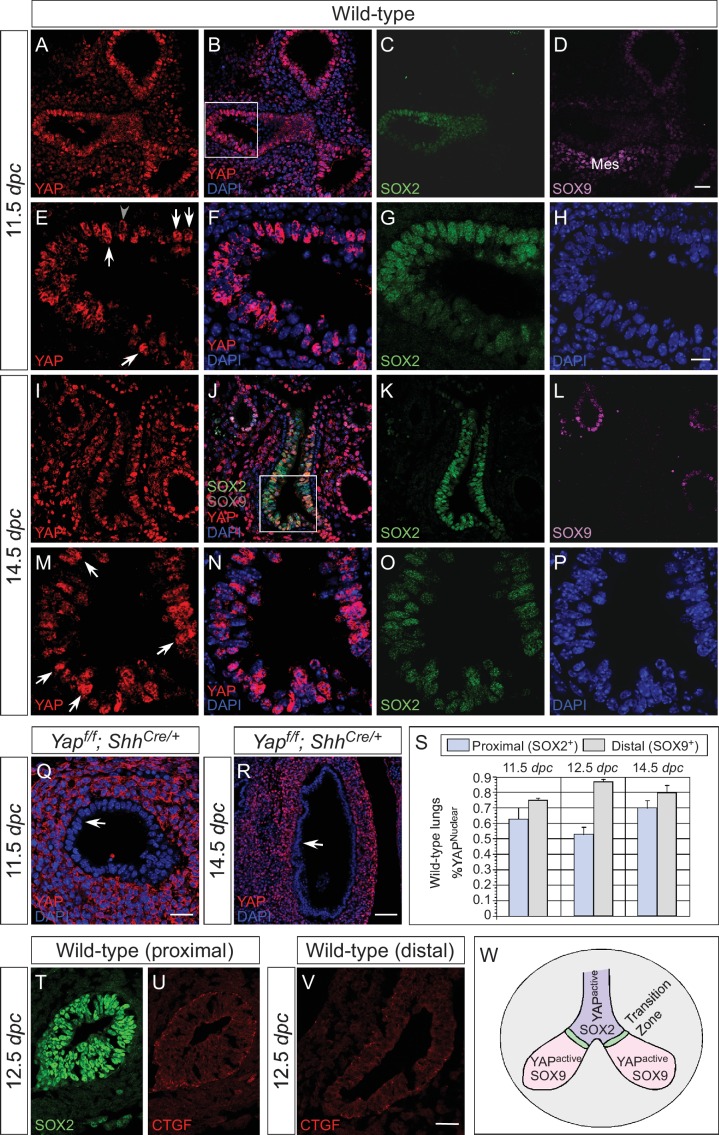
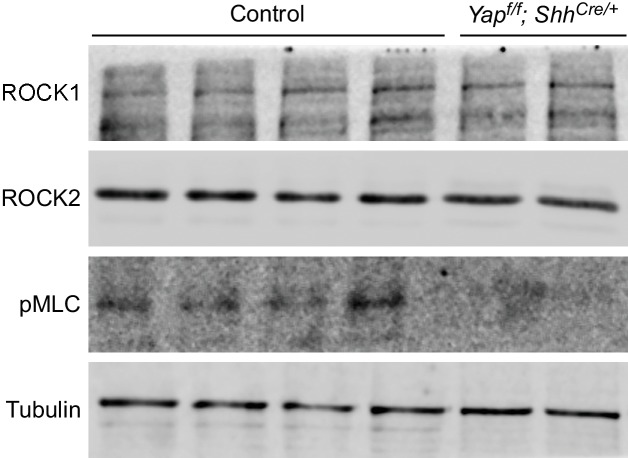
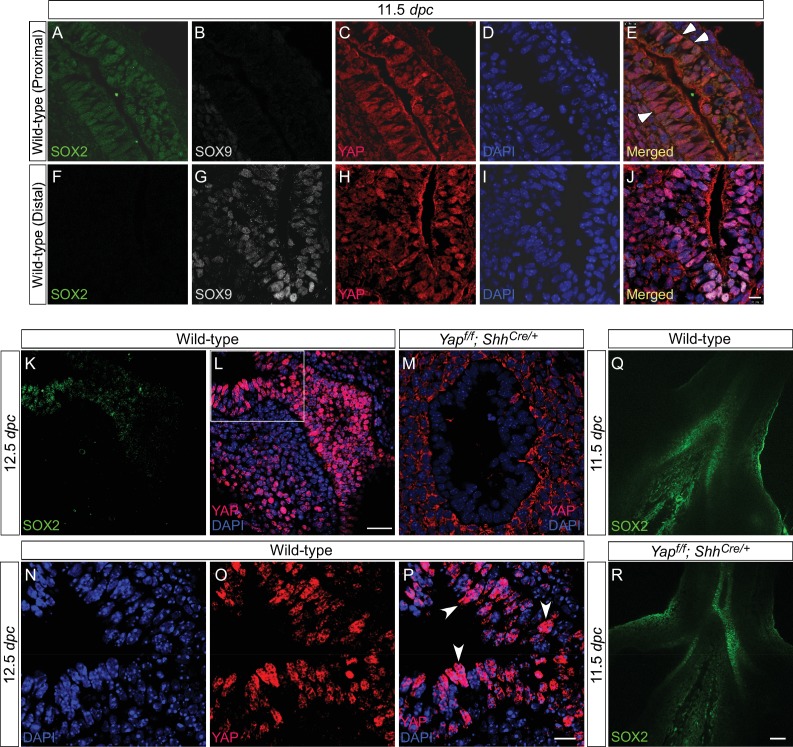
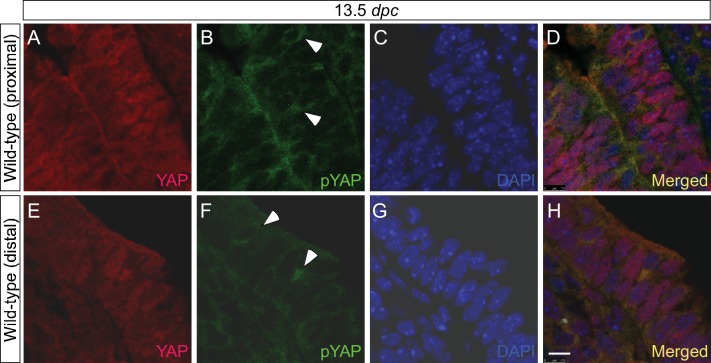
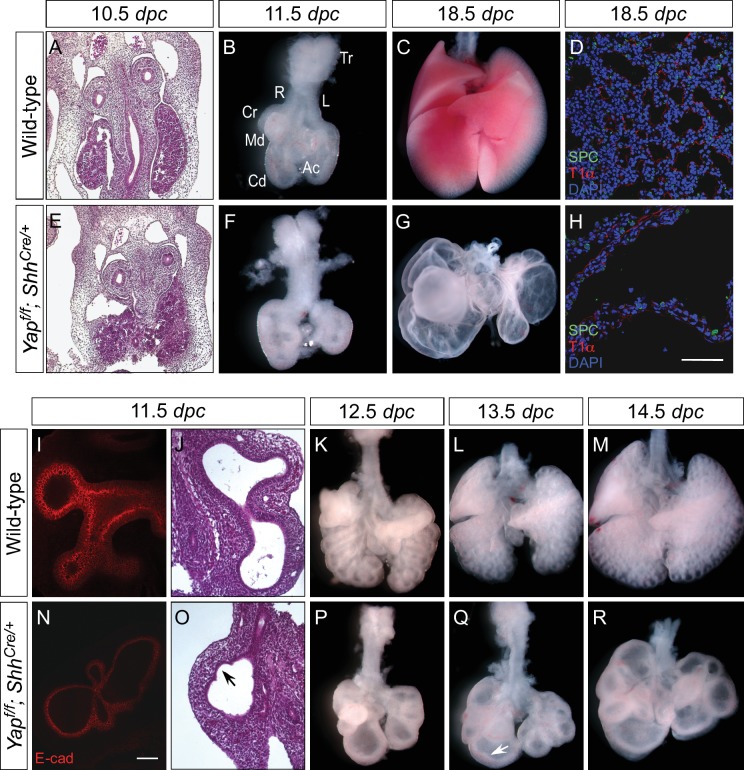
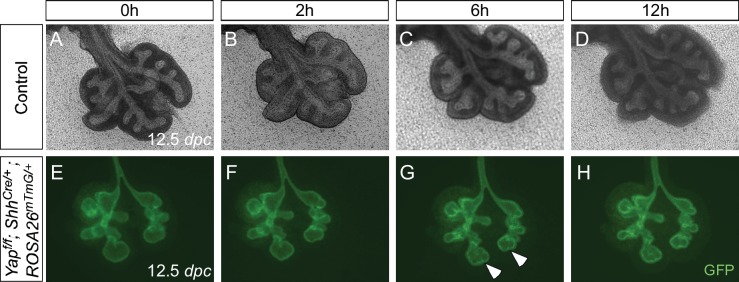
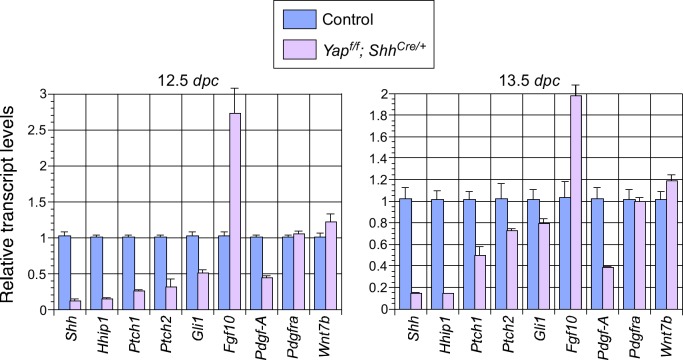
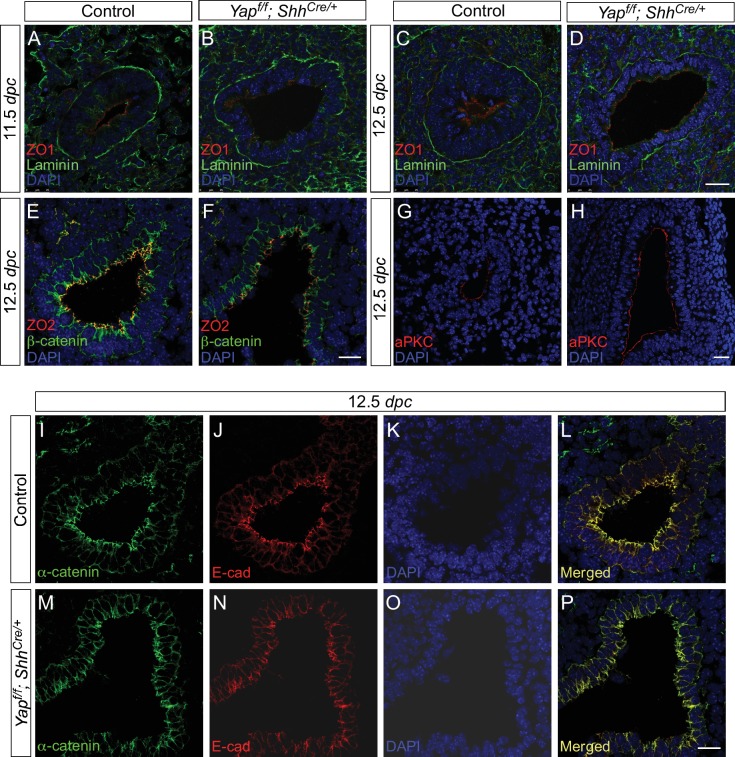
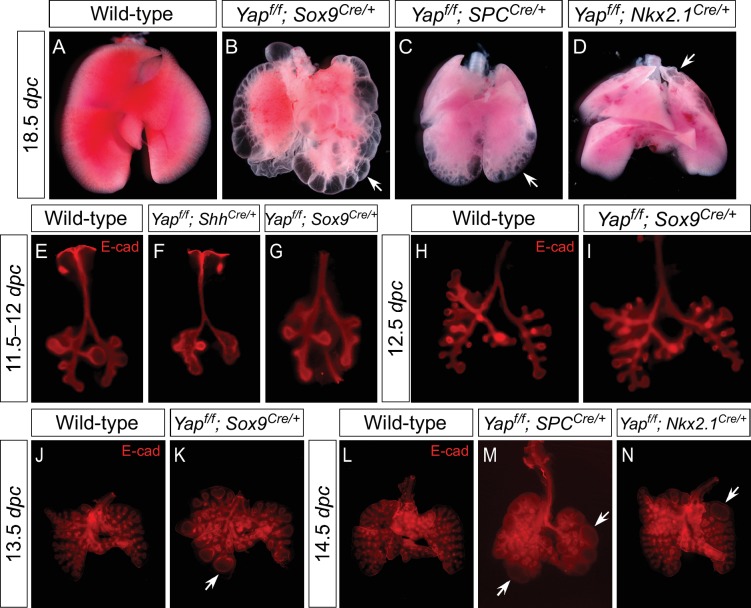
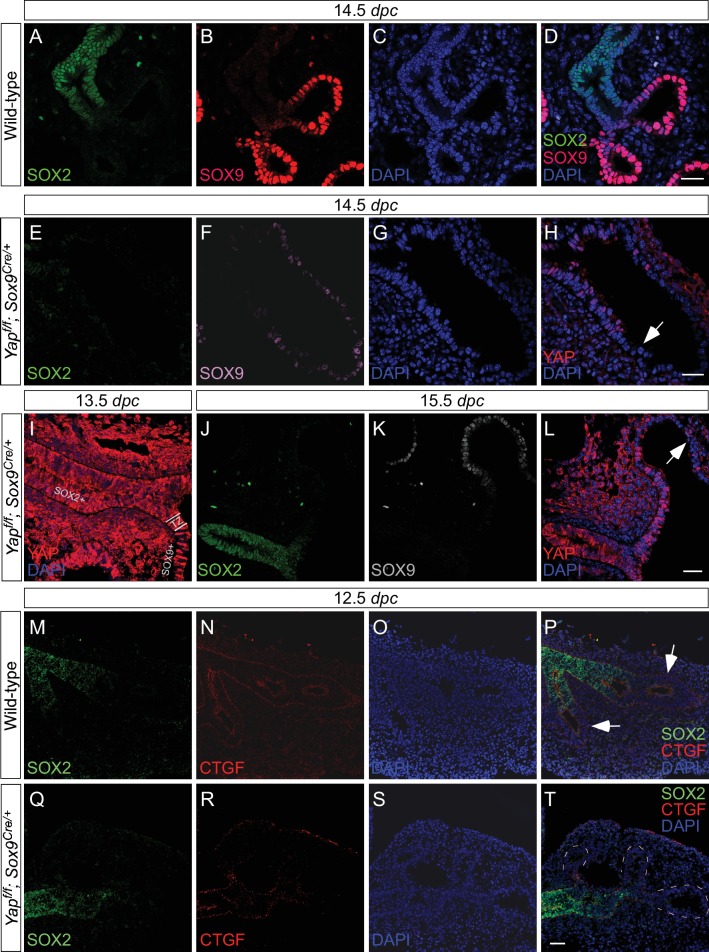
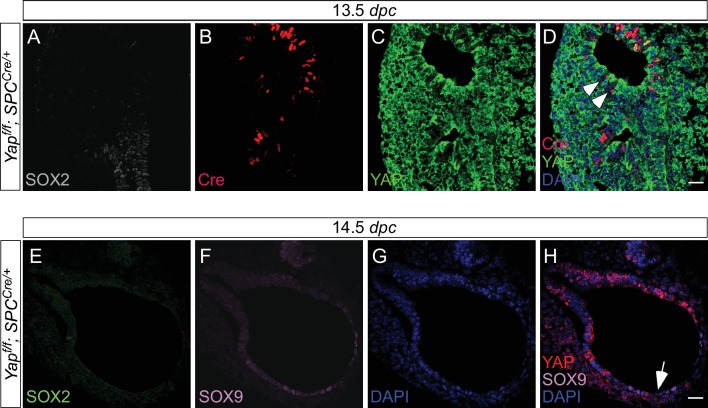
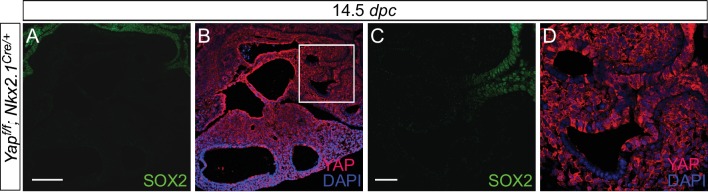
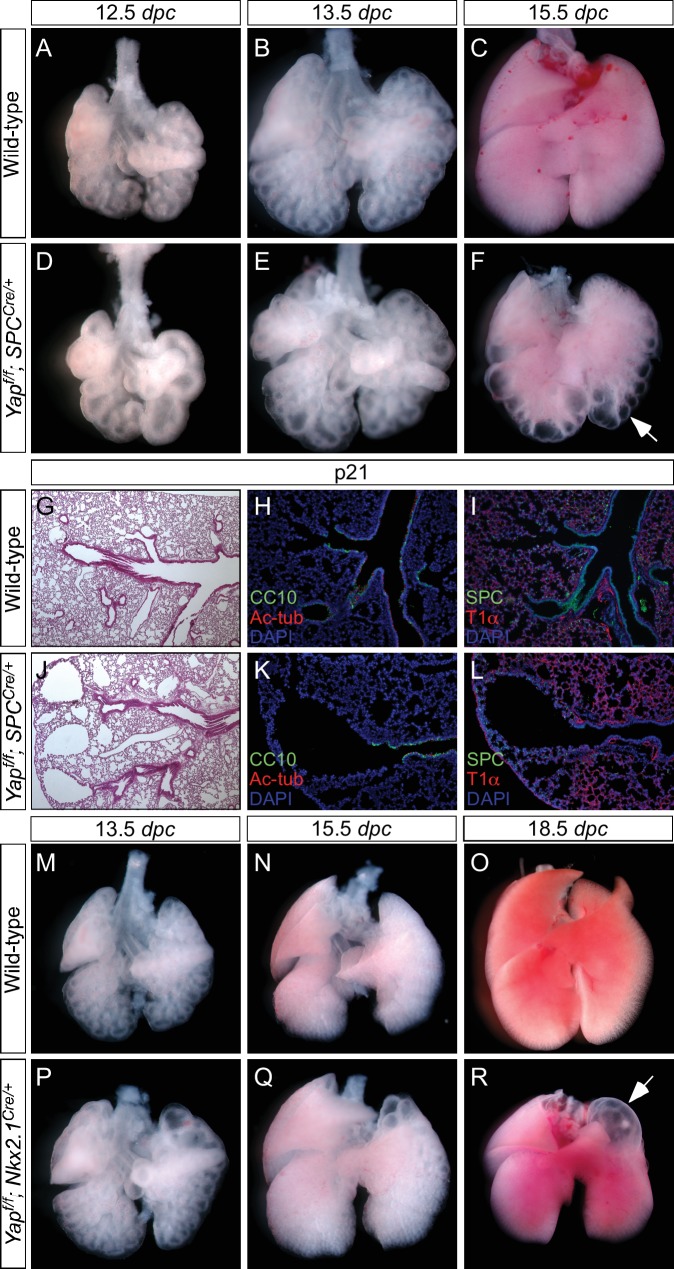
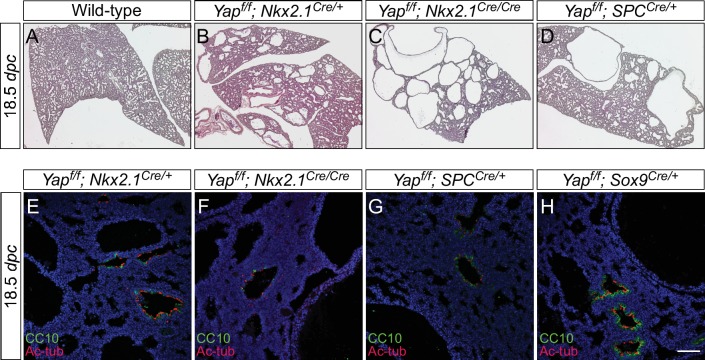
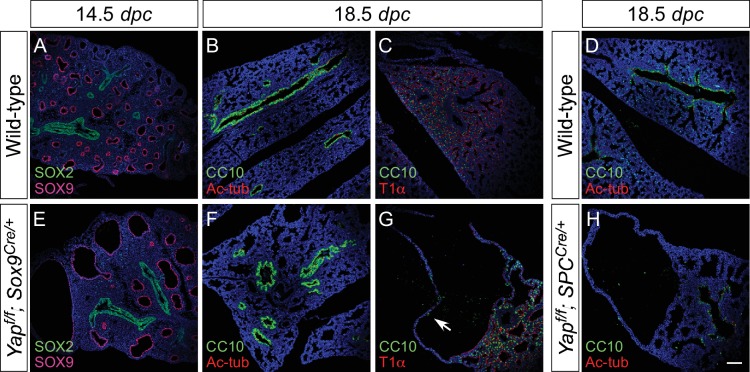
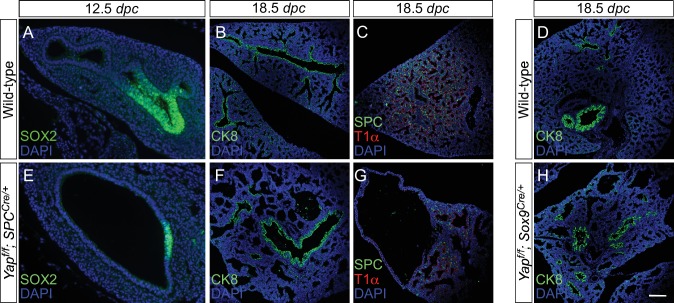
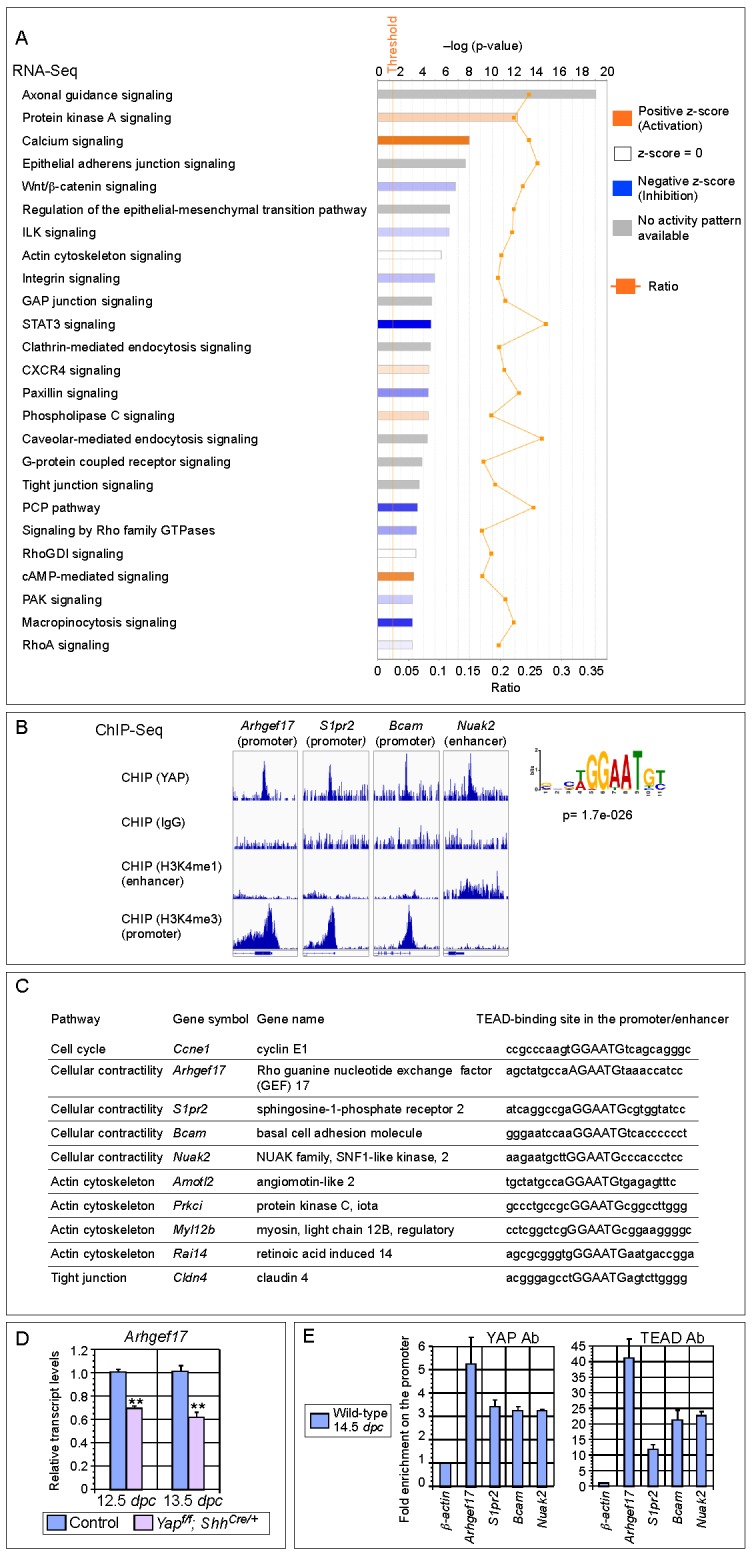
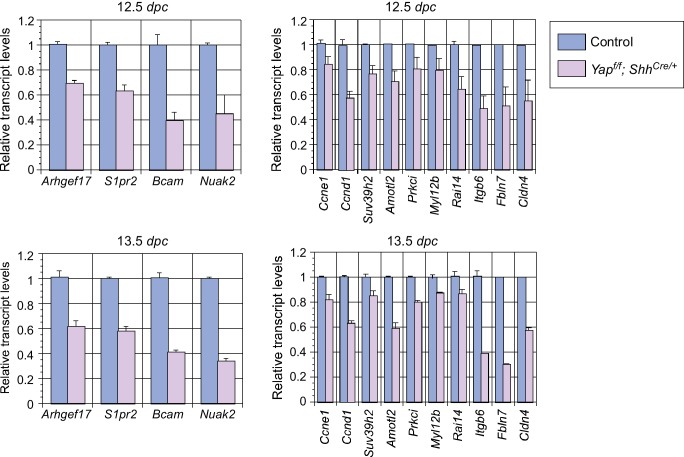
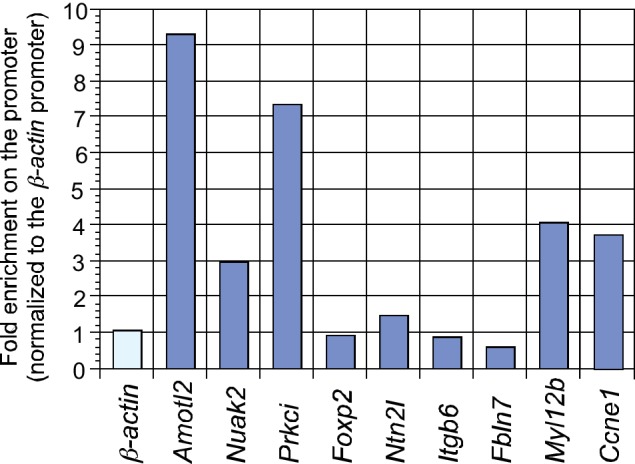
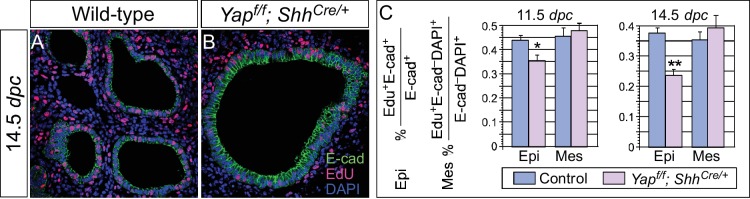
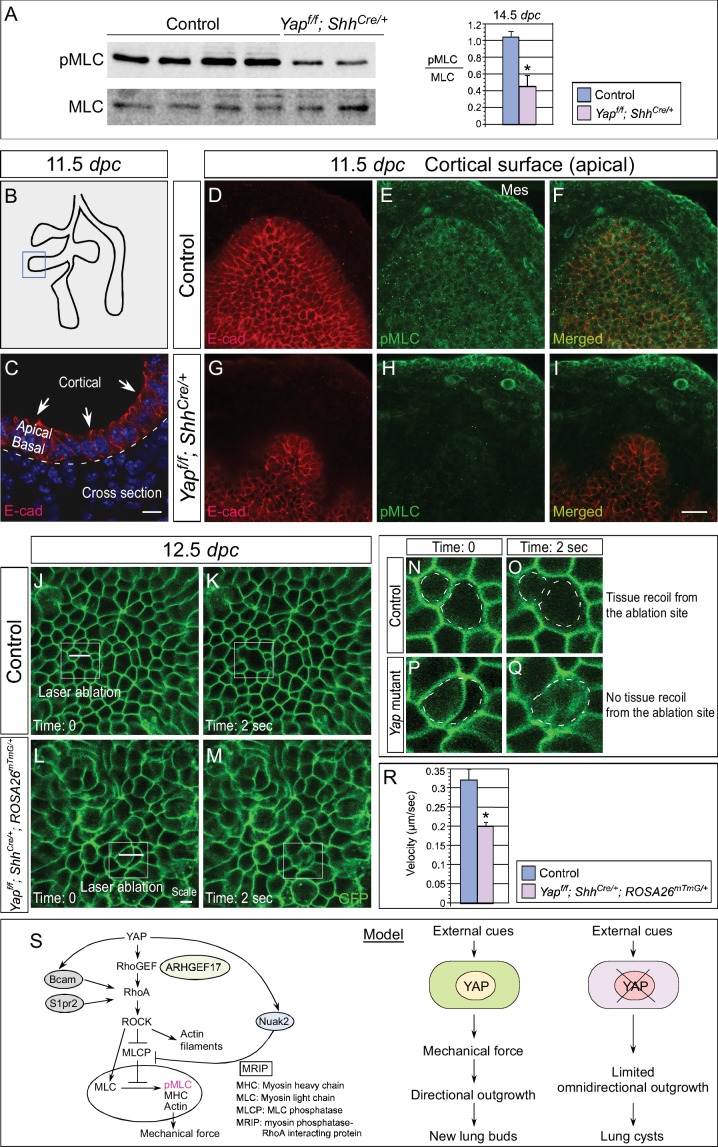
Branching morphogenesis is a fundamental program for tissue patterning. We show that active YAP, a key mediator of Hippo signaling, is distributed throughout the murine lung epithelium and loss of epithelial YAP severely disrupts branching. Failure to branch is restricted to regions where YAP activity is removed. This suggests that YAP controls local epithelial cell properties. In support of this model, mechanical force production is compromised and cell proliferation is reduced in mutant lungs. We propose that defective force generation and insufficient epithelial cell number underlie the branching defects. Through genomic analysis, we also uncovered a feedback control of pMLC levels, which is critical for mechanical force production, likely through the direct induction of multiple regulators by YAP. Our work provides a molecular pathway that could control epithelial cell properties required for proper morphogenetic movement and pattern formation.
分支形态发生是组织图案化的基本程序。我们发现,作为Hippo信号通路的关键介质,活性YAP分布于整个小鼠肺上皮,上皮YAP的缺失会严重破坏分支。分支失败仅限于去除YAP活性的区域。这表明YAP控制局部上皮细胞特性。支持该模型的是,突变肺中机械力产生受损且细胞增殖减少。我们提出,力产生缺陷和上皮细胞数量不足是分支缺陷的基础。通过基因组分析,我们还发现了对pMLC水平的反馈控制,这对机械力产生至关重要,可能是通过YAP直接诱导多种调节因子实现的。我们的工作提供了一条分子途径,可控制适当形态发生运动和模式形成所需的上皮细胞特性。