Dorrell Richard G, Gile Gillian, McCallum Giselle, Méheust Raphaël, Bapteste Eric P, Klinger Christen M, Brillet-Guéguen Loraine, Freeman Katalina D, Richter Daniel J, Bowler Chris
IBENS, Département de Biologie, École Normale Supérieure, CNRS, Inserm, PSL Research University, Paris, France.
School of Life Sciences, Arizona State University, Tempe, United States.
Elife. 2017 May 12;6:e23717. doi: 10.7554/eLife.23717.
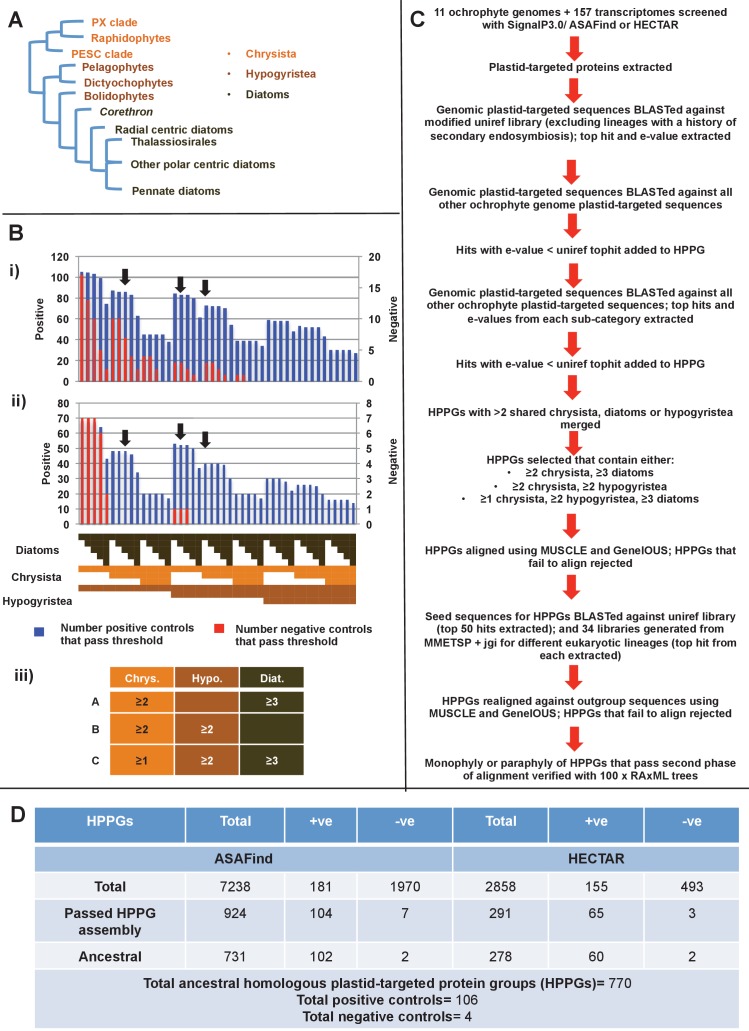
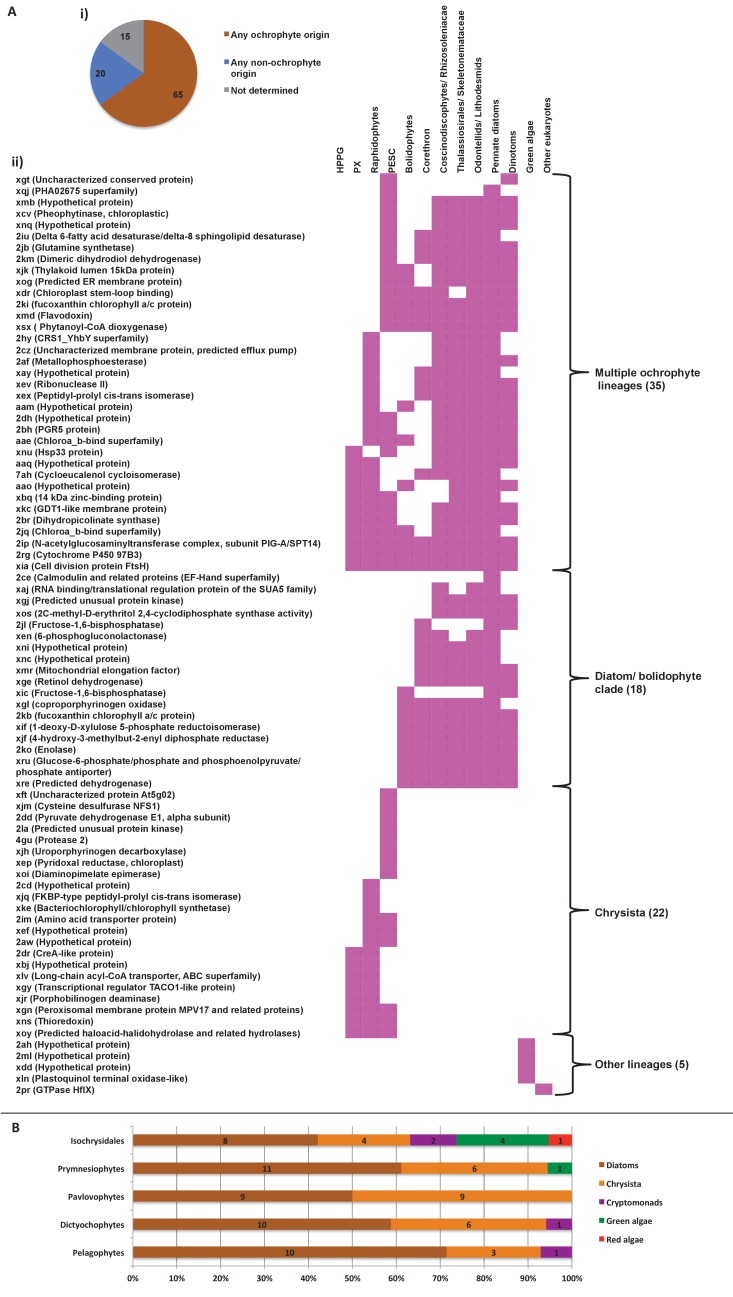
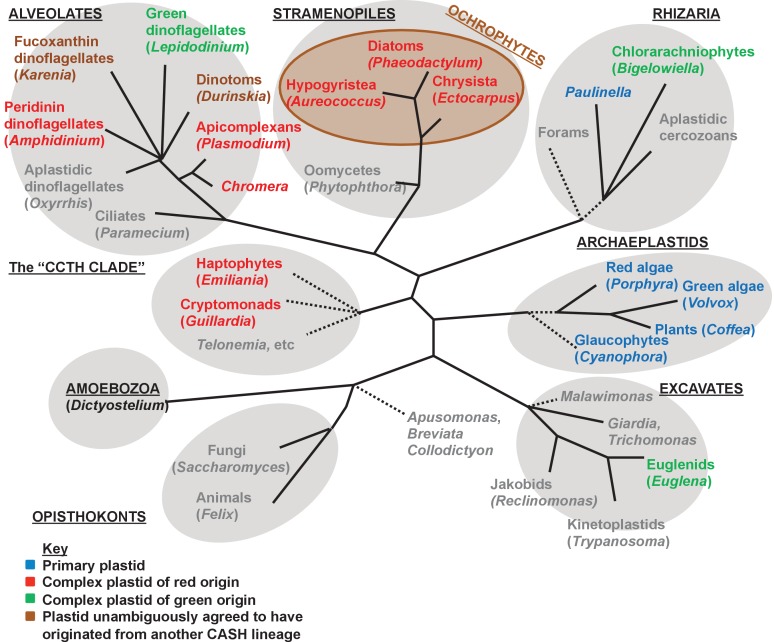
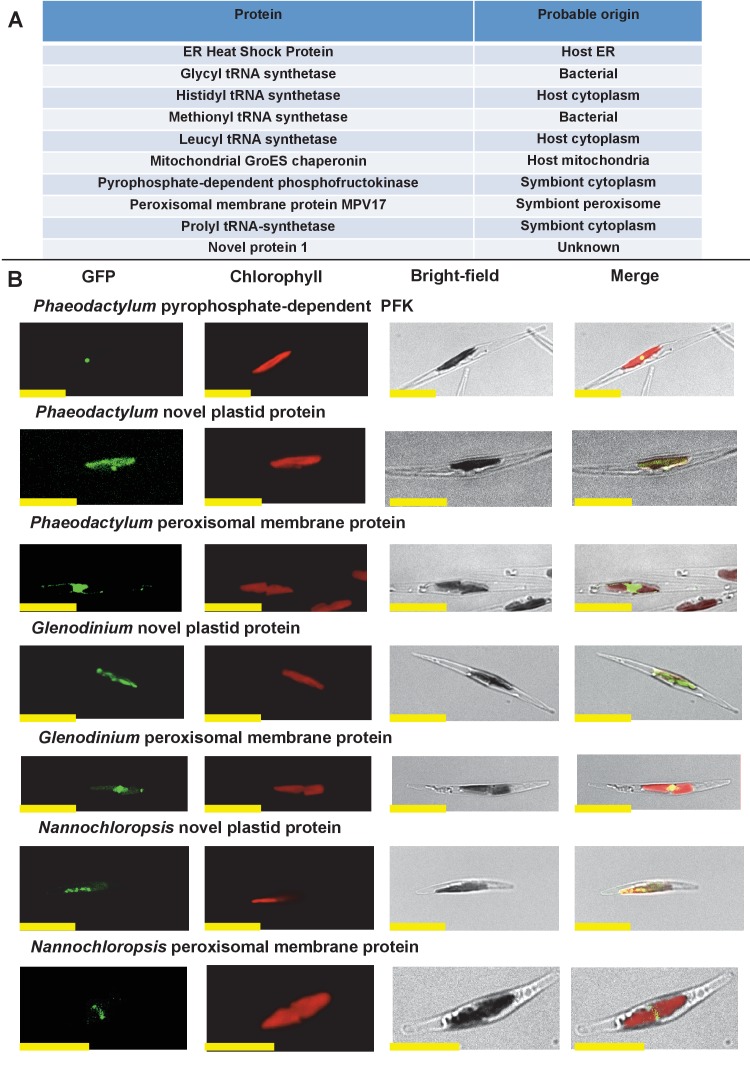
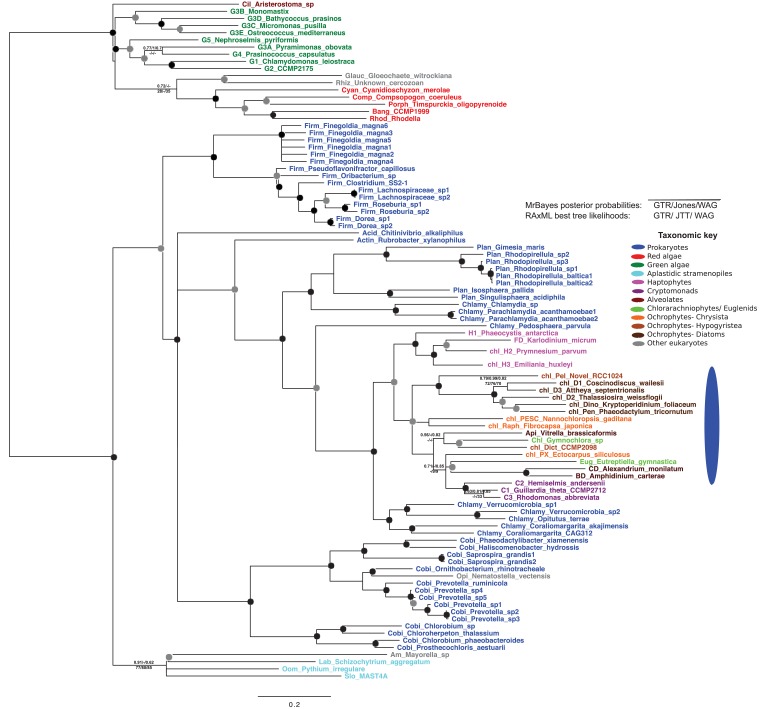
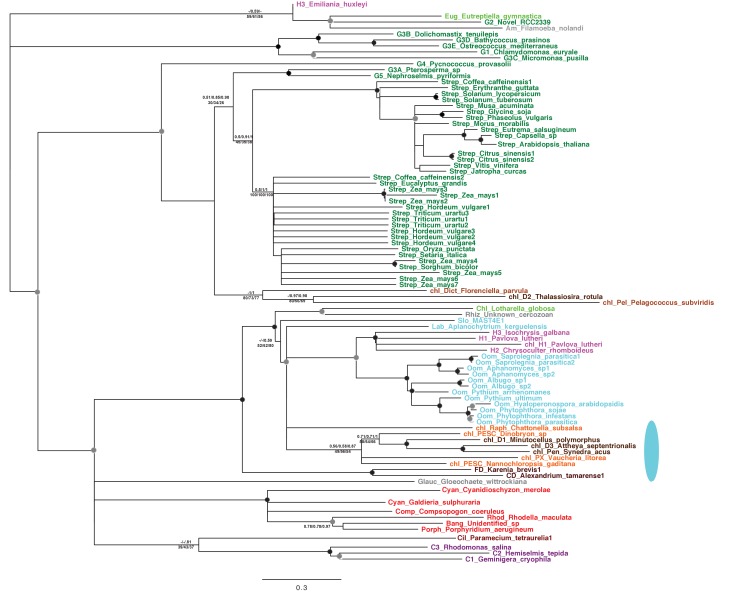
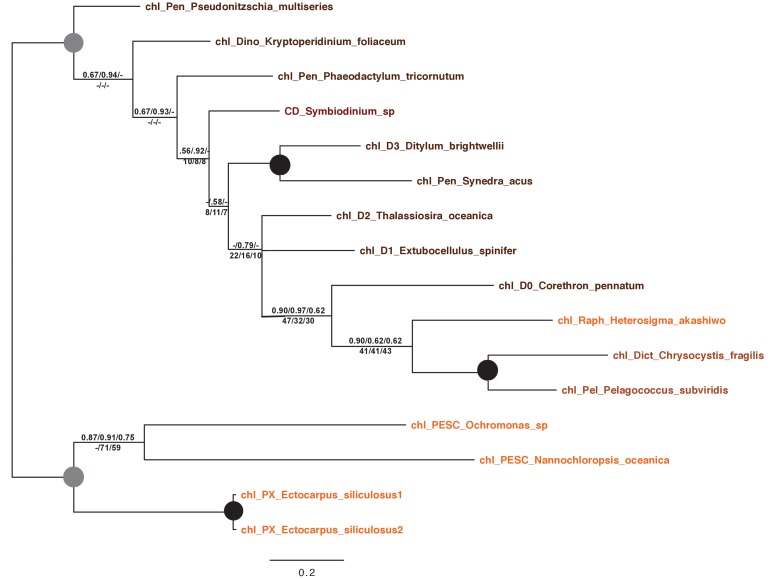
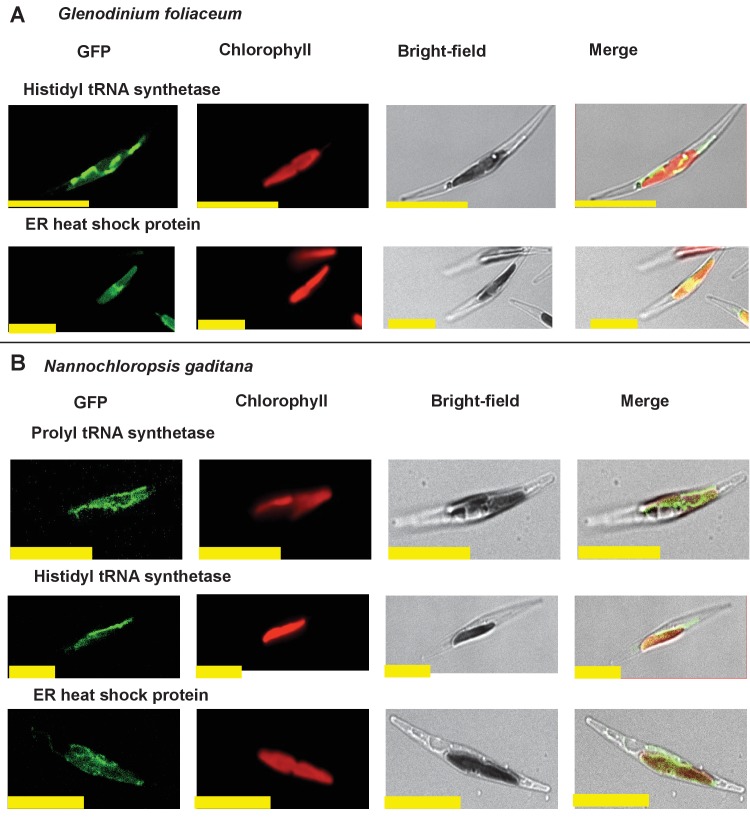
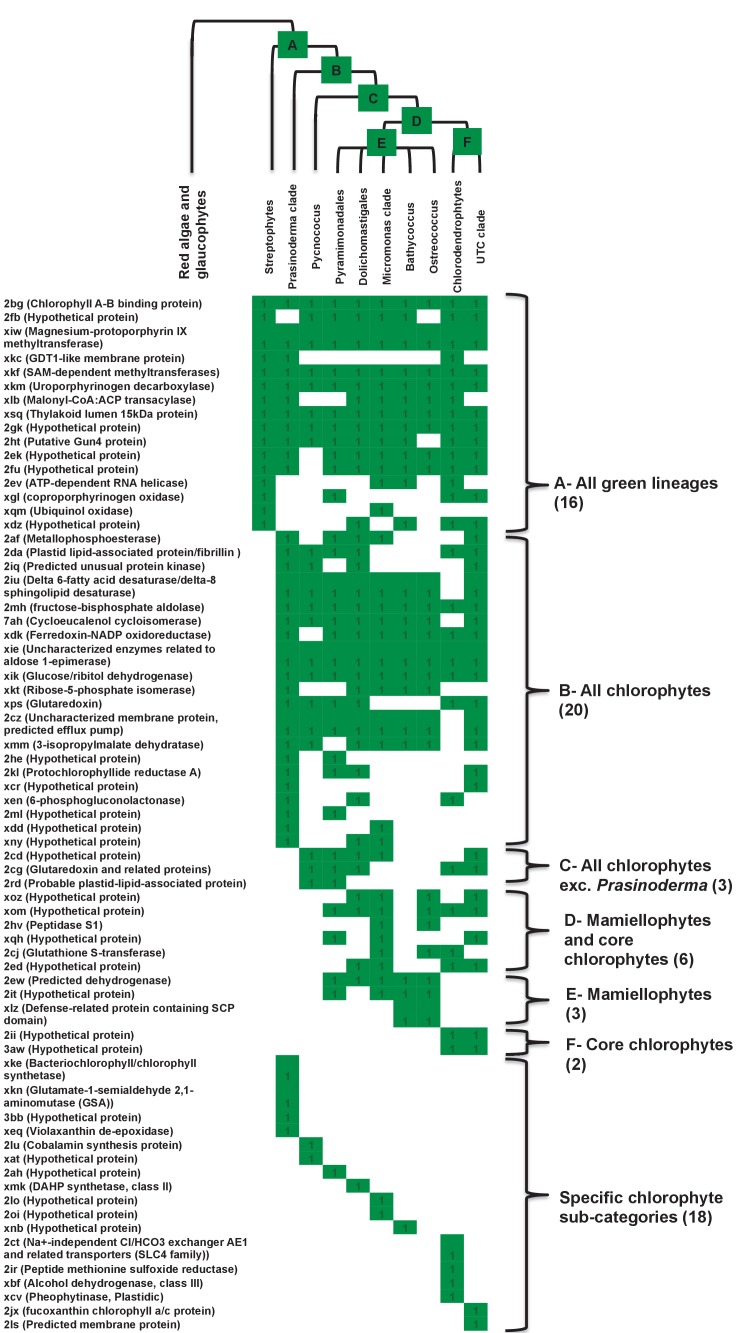
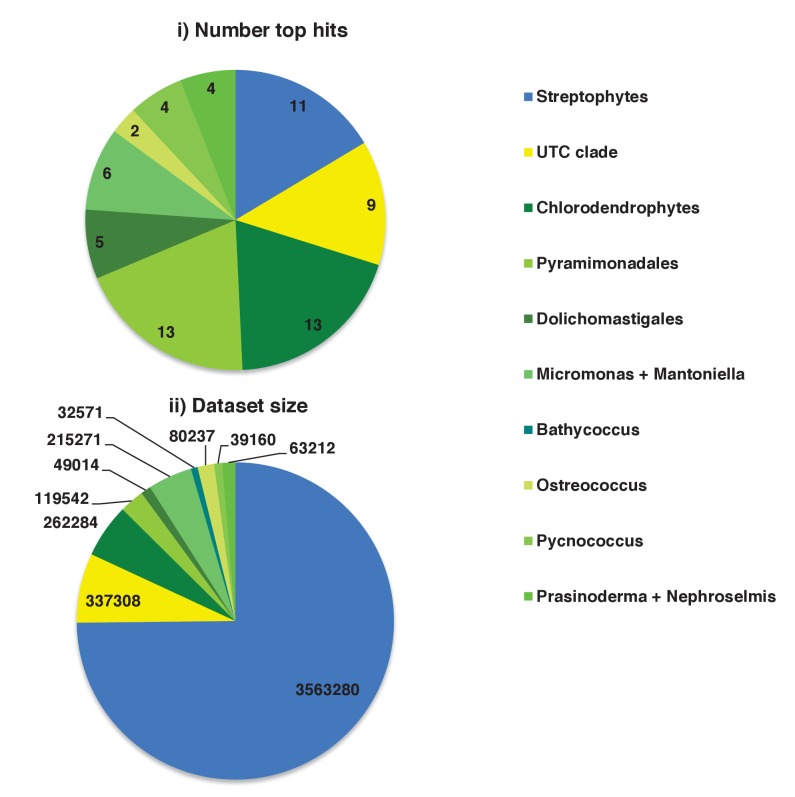
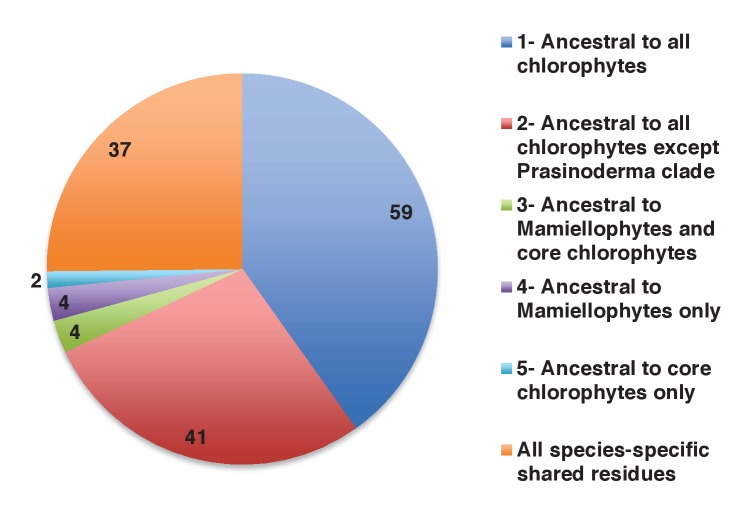
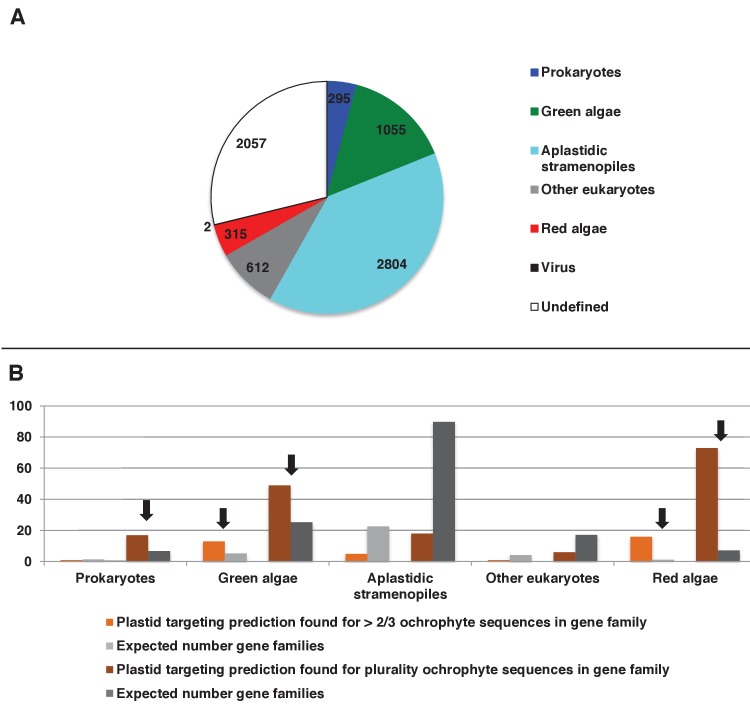
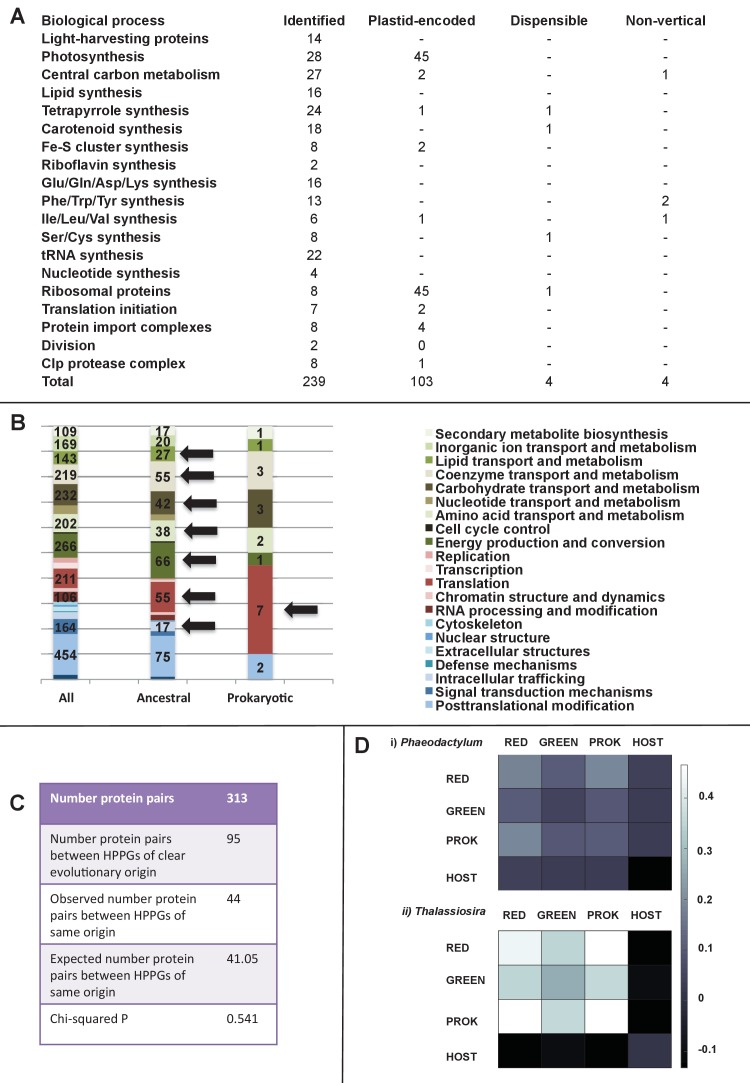
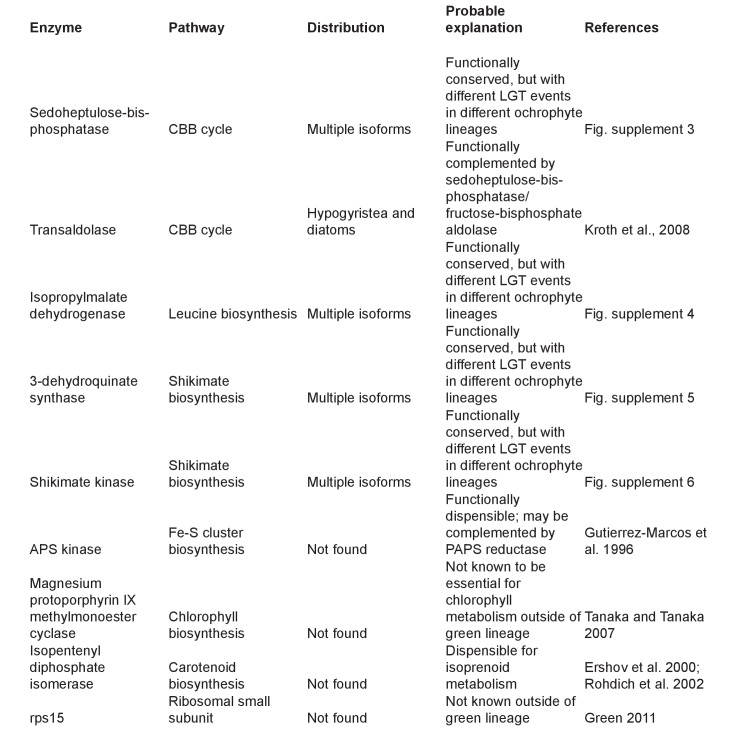
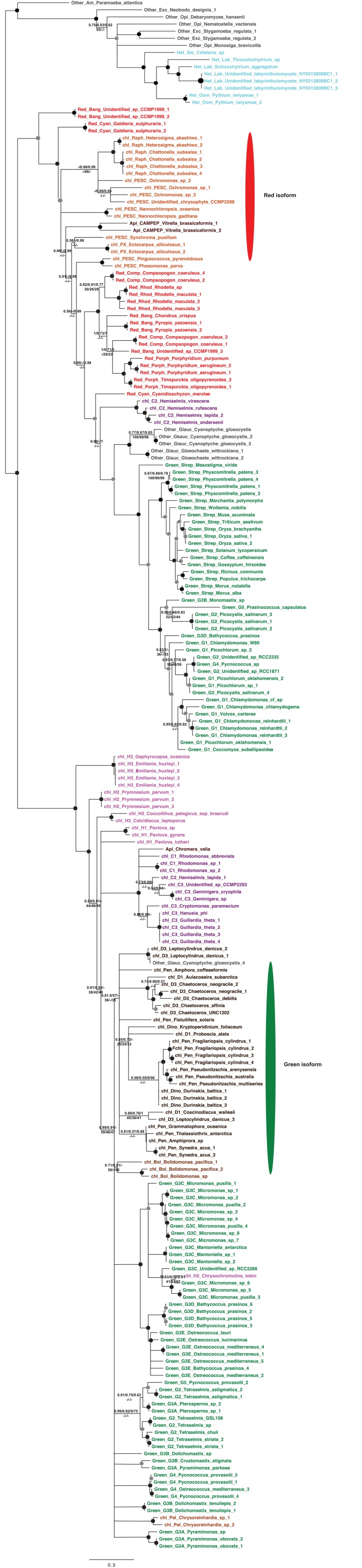
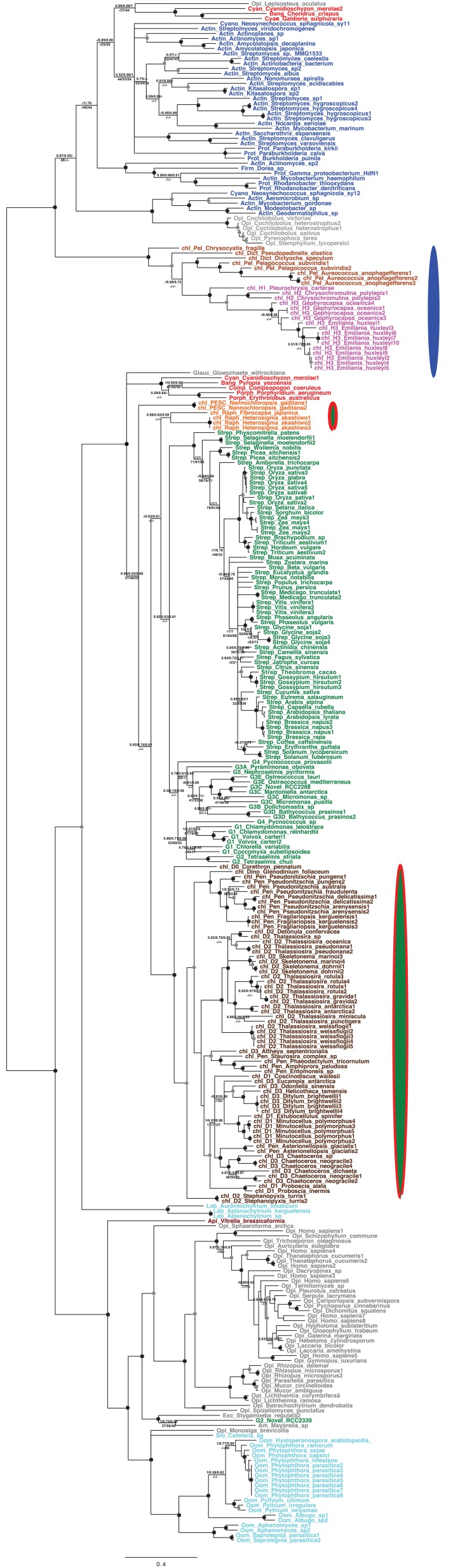
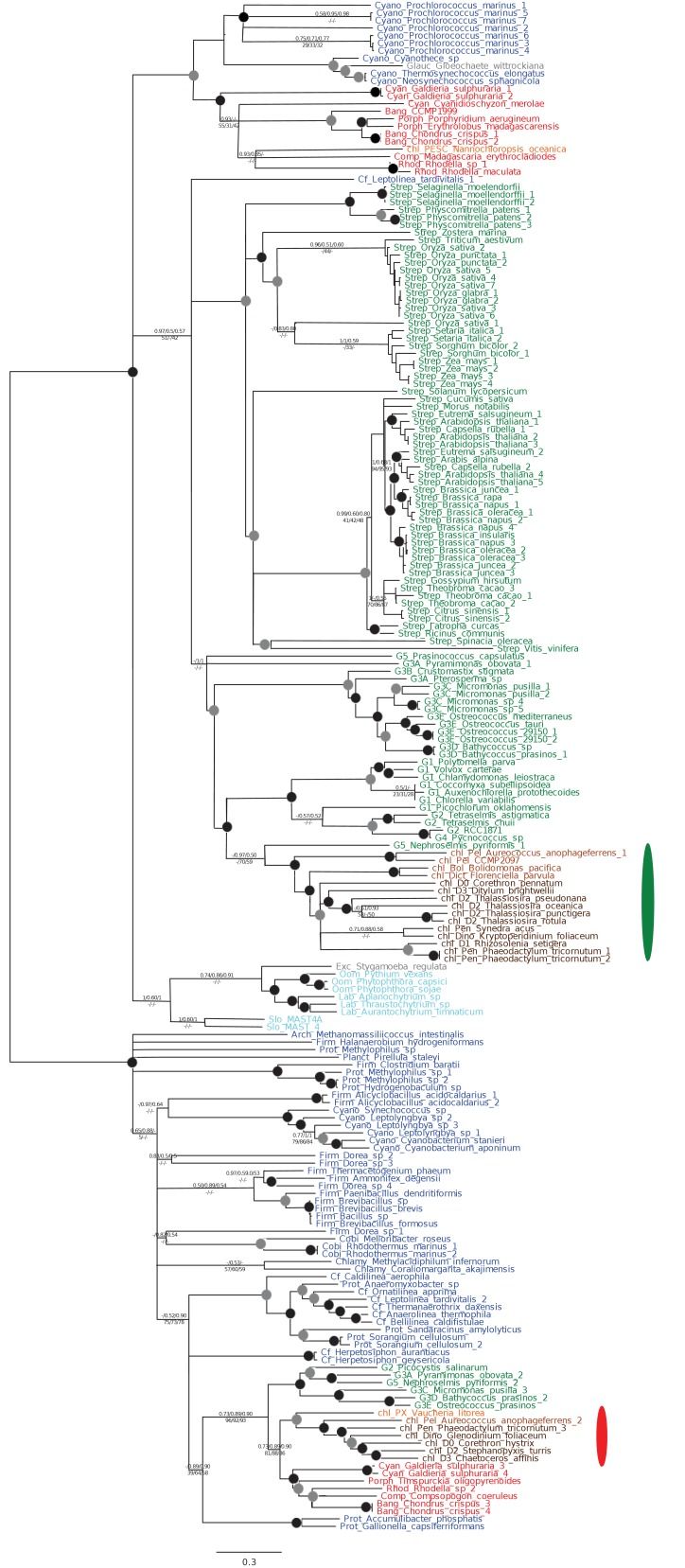
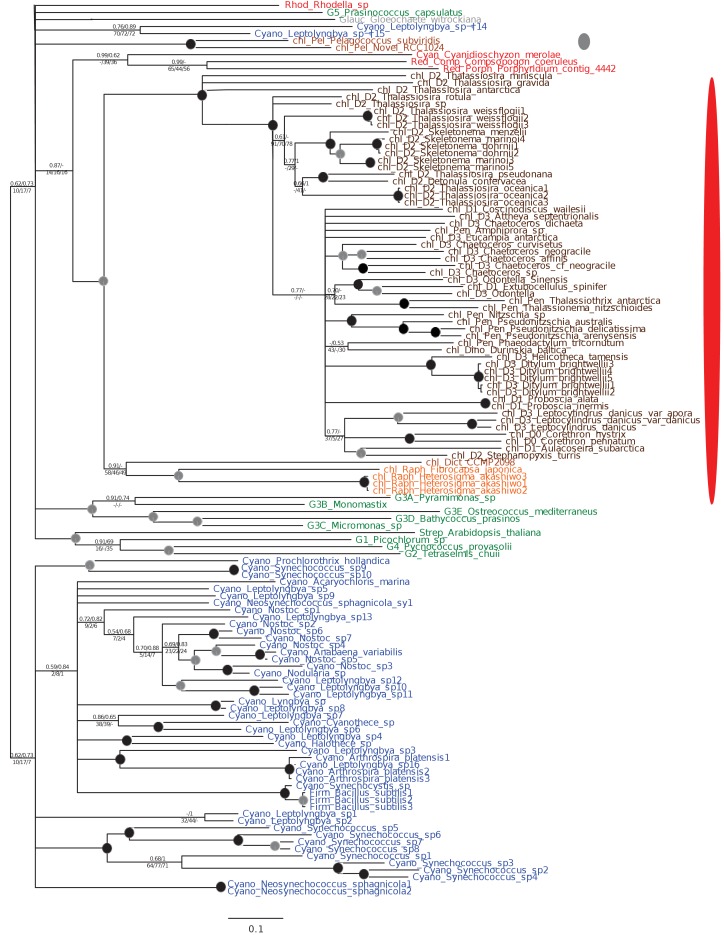
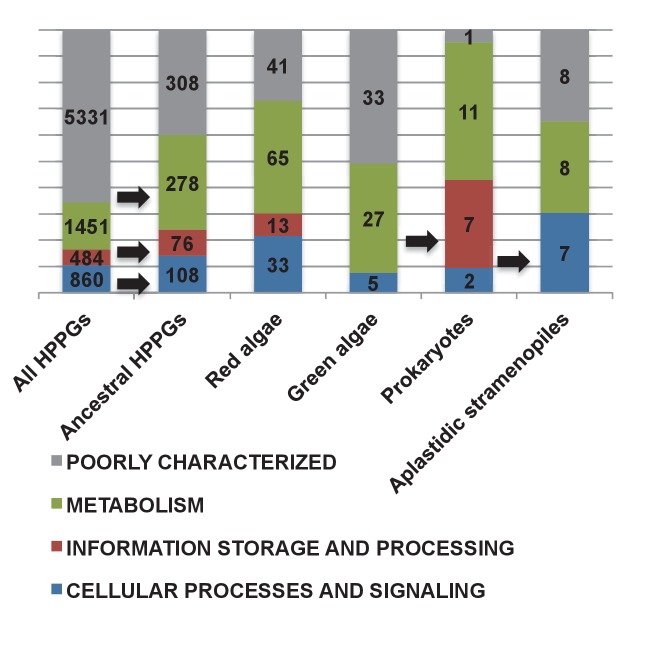
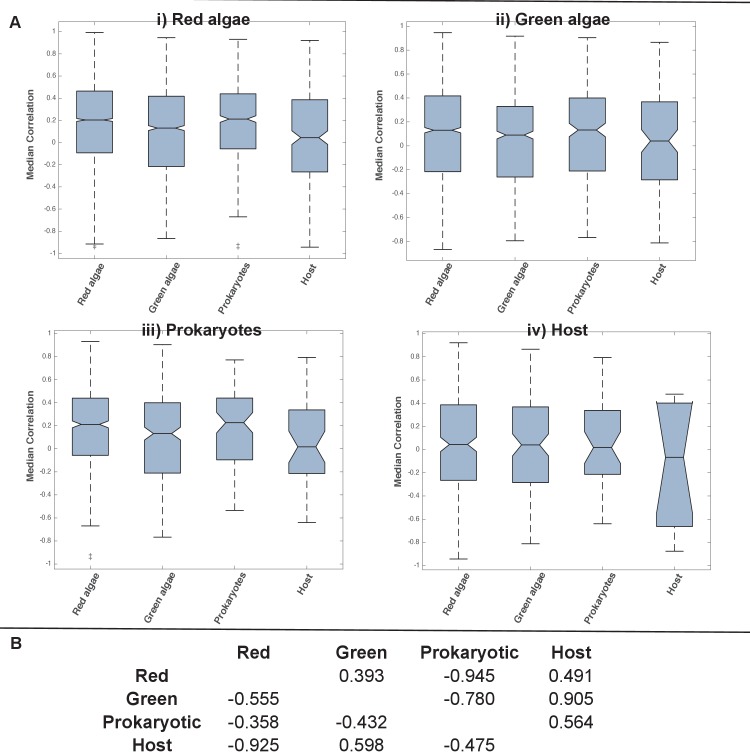
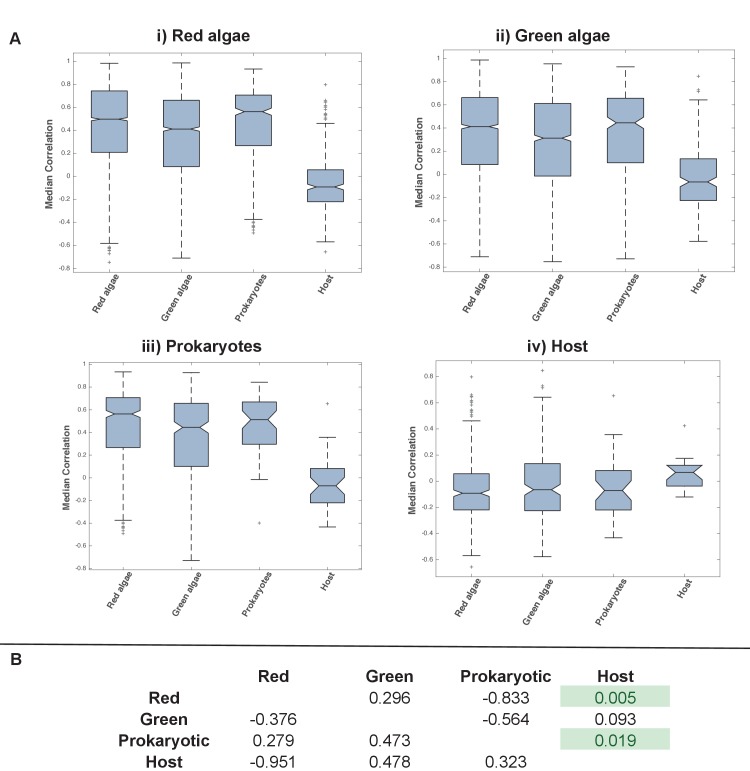
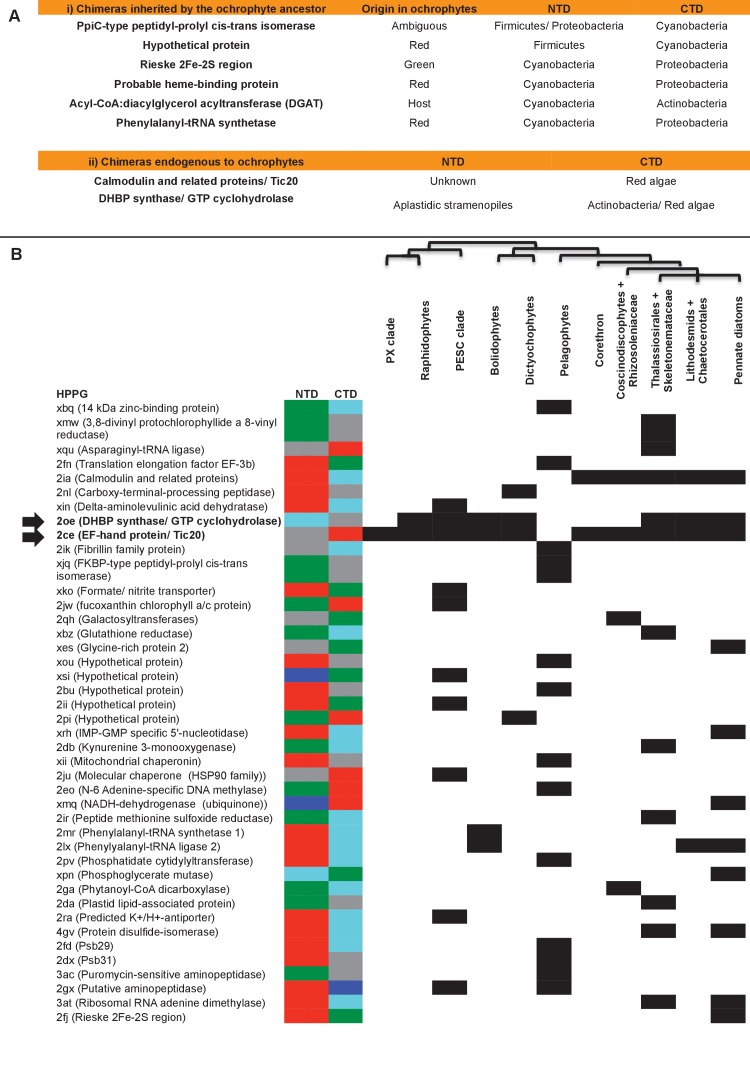
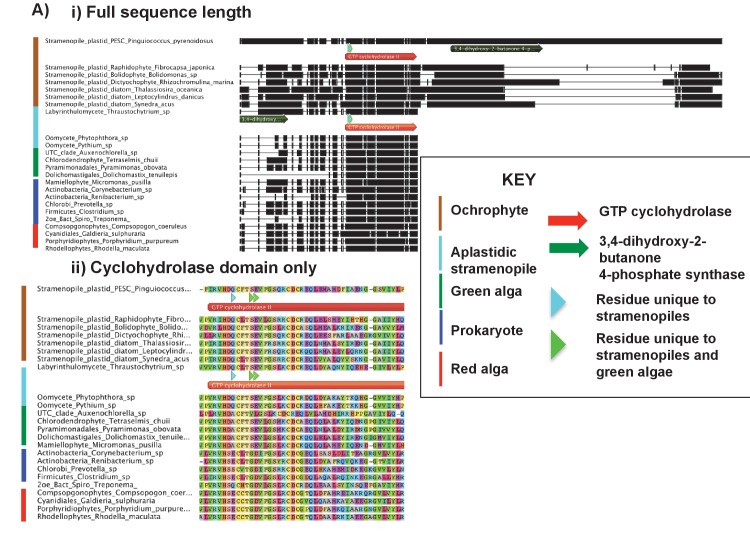
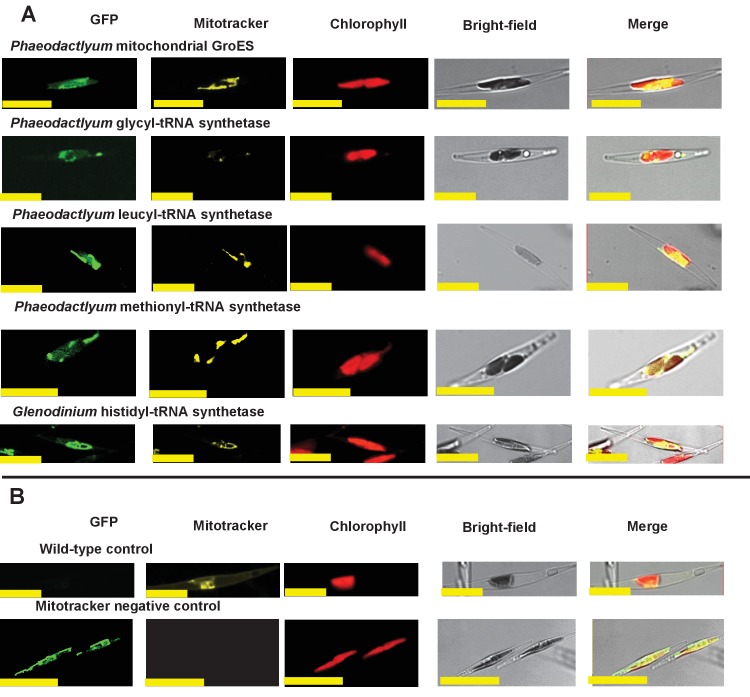
Plastids are supported by a wide range of proteins encoded within the nucleus and imported from the cytoplasm. These plastid-targeted proteins may originate from the endosymbiont, the host, or other sources entirely. Here, we identify and characterise 770 plastid-targeted proteins that are conserved across the ochrophytes, a major group of algae including diatoms, pelagophytes and kelps, that possess plastids derived from red algae. We show that the ancestral ochrophyte plastid proteome was an evolutionary chimera, with 25% of its phylogenetically tractable nucleus-encoded proteins deriving from green algae. We additionally show that functional mixing of host and plastid proteomes, such as through dual-targeting, is an ancestral feature of plastid evolution. Finally, we detect a clear phylogenetic signal from one ochrophyte subgroup, the lineage containing pelagophytes and dictyochophytes, in plastid-targeted proteins from another major algal lineage, the haptophytes. This may represent a possible serial endosymbiosis event deep in eukaryotic evolutionary history.
质体由细胞核内编码并从细胞质中导入的多种蛋白质支持。这些靶向质体的蛋白质可能完全来自内共生体、宿主或其他来源。在这里,我们鉴定并表征了770种靶向质体的蛋白质,它们在褐藻植物中是保守的,褐藻植物是包括硅藻、褐囊藻和海带在内的一大类藻类,它们拥有源自红藻的质体。我们表明,原始褐藻植物质体蛋白质组是一种进化嵌合体,其系统发育可追踪的细胞核编码蛋白质中有25%来自绿藻。我们还表明,宿主和质体蛋白质组的功能混合,例如通过双靶向,是质体进化的一个原始特征。最后,我们在另一个主要藻类谱系——定鞭藻门的靶向质体蛋白质中,检测到来自褐藻植物的一个亚群(包含褐囊藻和网柱藻的谱系)的清晰系统发育信号。这可能代表了真核生物进化历史深处可能发生的一系列内共生事件。