Fong Nova, Saldi Tassa, Sheridan Ryan M, Cortazar Michael A, Bentley David L
Department of Biochemistry and Molecular Genetics, University of Colorado School of Medicine, PO Box 6511, Aurora, CO 80045, USA.
Department of Biochemistry and Molecular Genetics, University of Colorado School of Medicine, PO Box 6511, Aurora, CO 80045, USA.
Mol Cell. 2017 May 18;66(4):546-557.e3. doi: 10.1016/j.molcel.2017.04.016. Epub 2017 May 11.
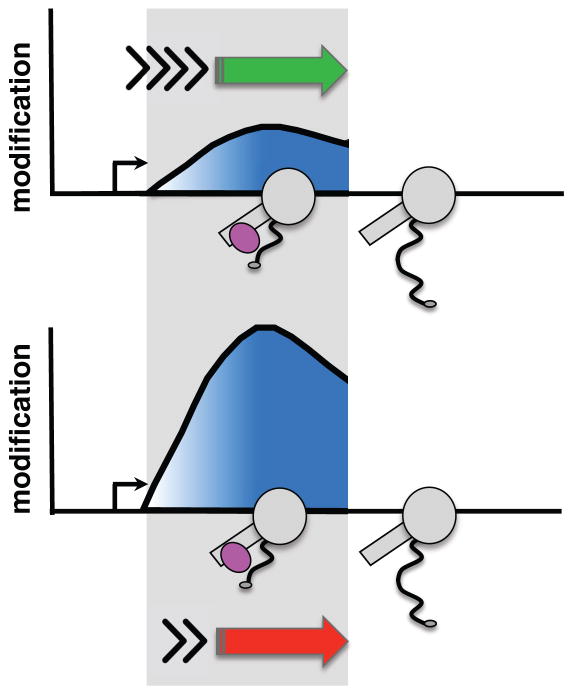
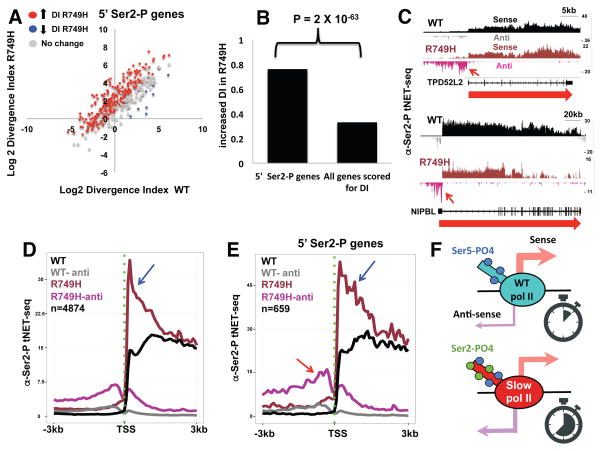
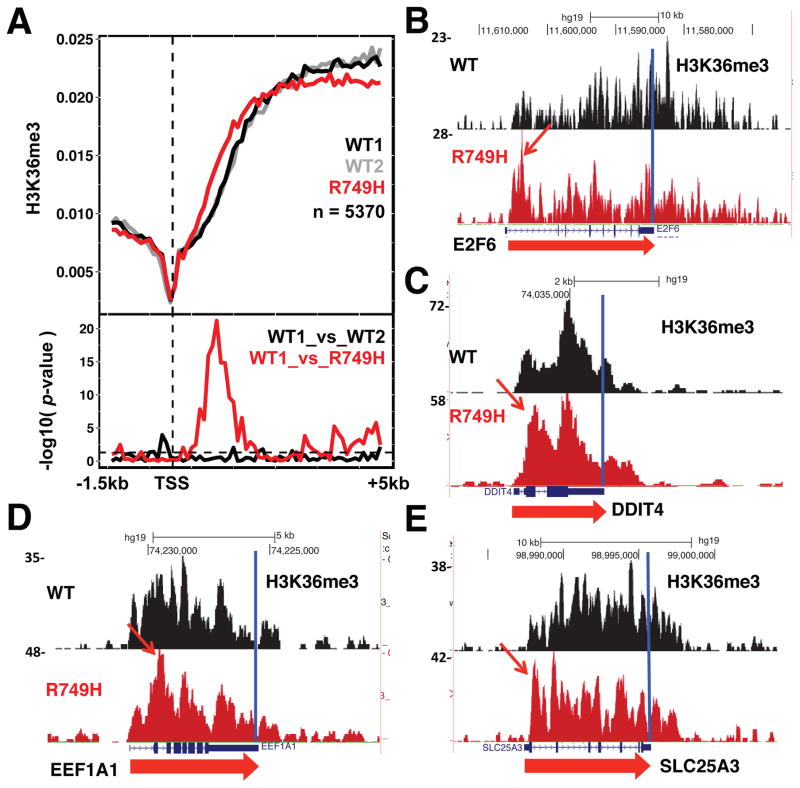
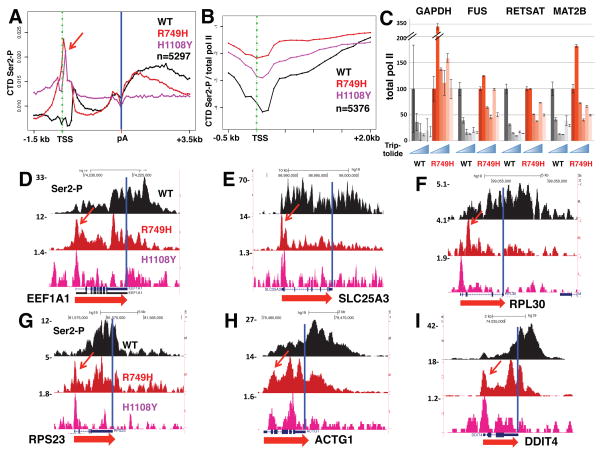
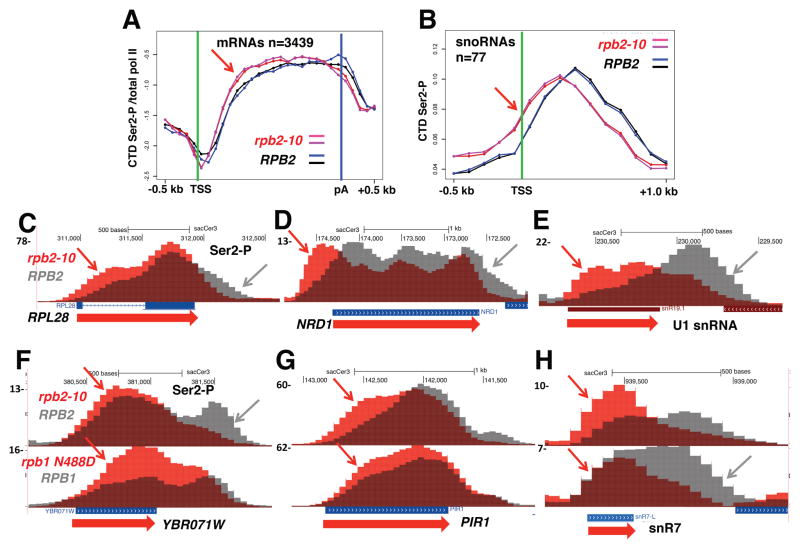
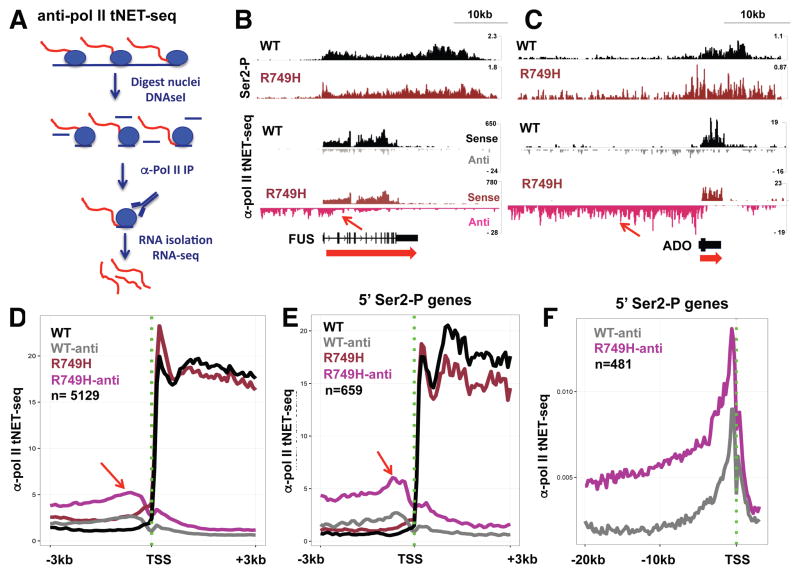
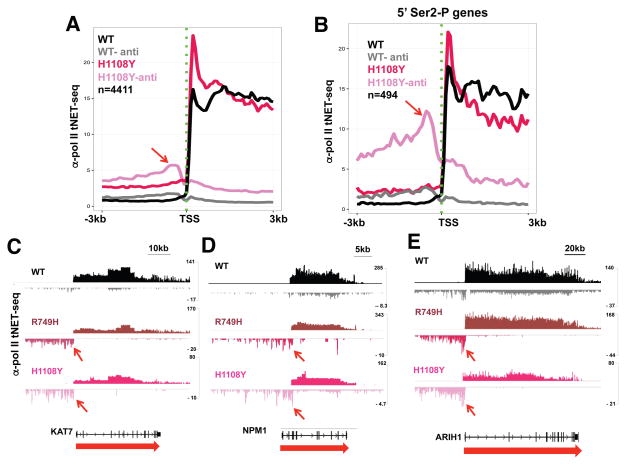
Eukaryotic genes are marked by conserved post-translational modifications on the RNA pol II C-terminal domain (CTD) and the chromatin template. How the 5'-3' profiles of these marks are established is poorly understood. Using pol II mutants in human cells, we found that slow transcription repositioned specific co-transcriptionally deposited chromatin modifications; histone H3 lysine 36 trimethyl (H3K36me3) shifted within genes toward 5' ends, and histone H3 lysine 4 dimethyl (H3K4me2) extended farther upstream of start sites. Slow transcription also evoked a hyperphosphorylation of CTD Ser2 residues at 5' ends of genes that is conserved in yeast. We propose a "dwell time in the target zone" model to explain the effects of transcriptional dynamics on the establishment of co-transcriptionally deposited protein modifications. Promoter-proximal Ser2 phosphorylation is associated with a longer pol II dwell time at start sites and reduced transcriptional polarity because of strongly enhanced divergent antisense transcription at promoters. These results demonstrate that pol II dynamics help govern the decision between sense and divergent antisense transcription.
真核基因由RNA聚合酶II羧基末端结构域(CTD)和染色质模板上保守的翻译后修饰标记。这些标记的5'-3'分布是如何建立的,目前还知之甚少。通过在人类细胞中使用聚合酶II突变体,我们发现缓慢转录会重新定位特定的共转录沉积染色质修饰;组蛋白H3赖氨酸36三甲基化(H3K36me3)在基因内朝着5'端移动,组蛋白H3赖氨酸4二甲基化(H3K4me2)在起始位点上游延伸得更远。缓慢转录还会引发基因5'端CTD丝氨酸2残基的过度磷酸化,这在酵母中是保守的。我们提出了一个“在靶区域的停留时间”模型来解释转录动力学对共转录沉积蛋白质修饰建立的影响。启动子近端丝氨酸2磷酸化与聚合酶II在起始位点的较长停留时间以及转录极性降低有关,这是由于启动子处强烈增强的反向反义转录所致。这些结果表明,聚合酶II动力学有助于控制有义转录和反向反义转录之间的抉择。