Fan Fushun, Matsunaga Kohichi, Wang Hao, Ishizaki Ray, Kobayashi Eri, Kiyonari Hiroshi, Mukumoto Yoshiko, Okunishi Katsuhide, Izumi Tetsuro
Laboratory of Molecular Endocrinology and Metabolism, Department of Molecular Medicine, Institute for Molecular and Cellular Regulation, Gunma University, Maebashi, Japan.
Animal Resource Development Unit, RIKEN Center for Life Science Technologies, Kobe, Japan.
Elife. 2017 Jul 4;6:e26174. doi: 10.7554/eLife.26174.
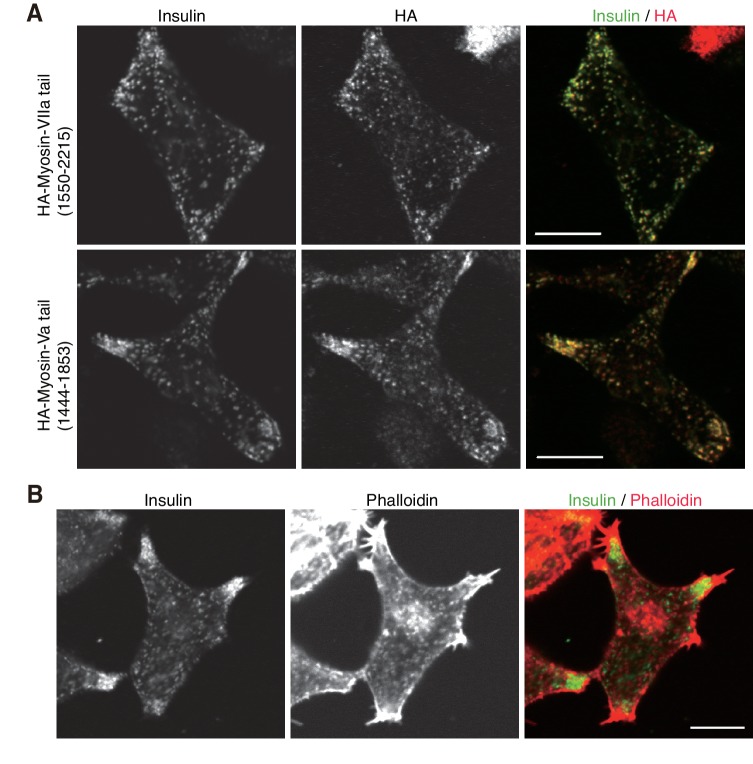
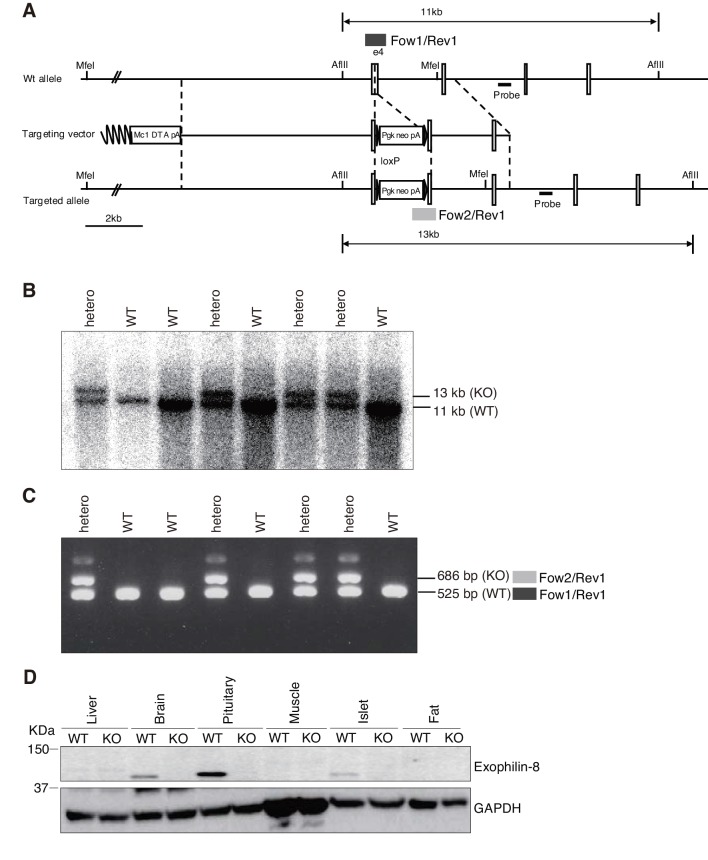
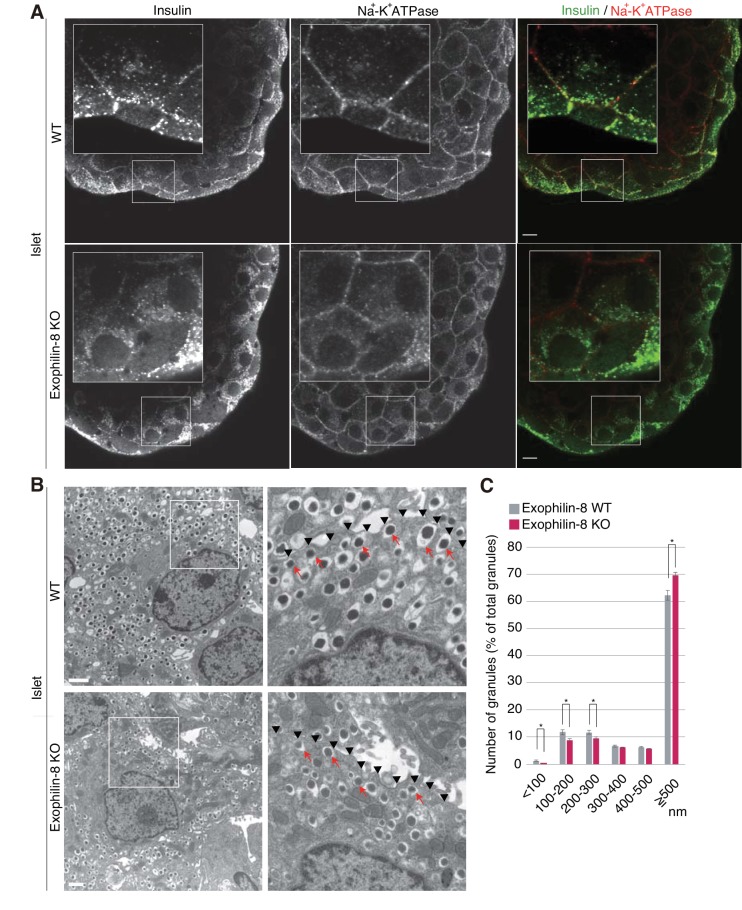
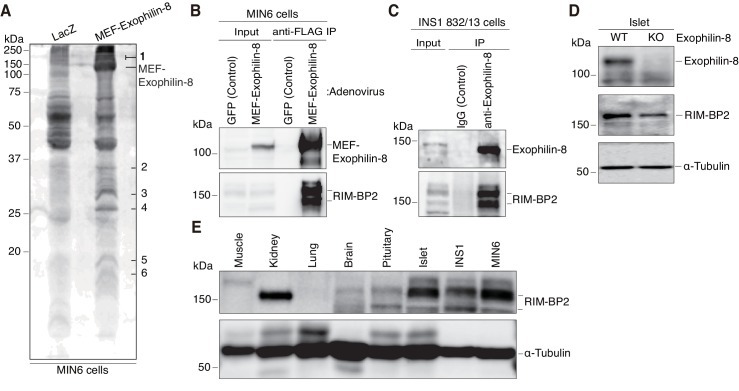
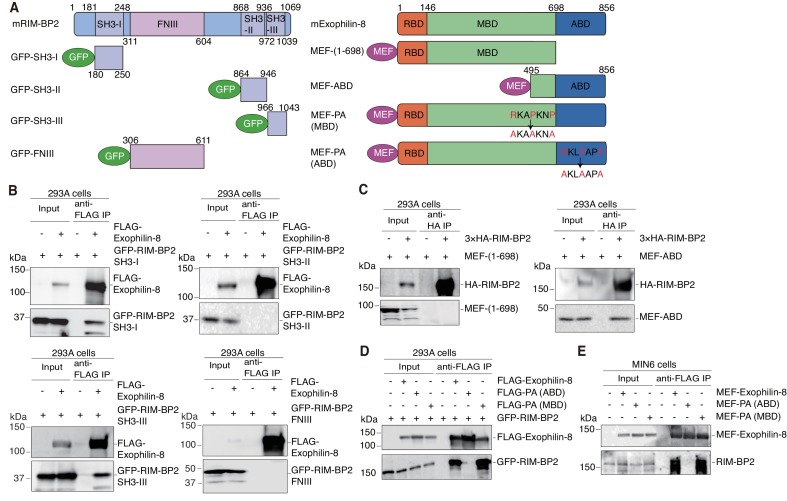
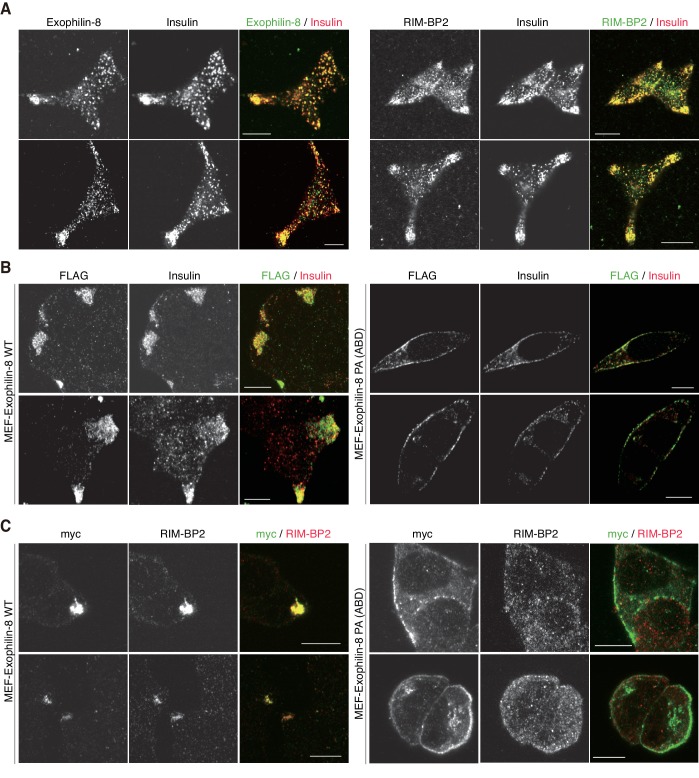
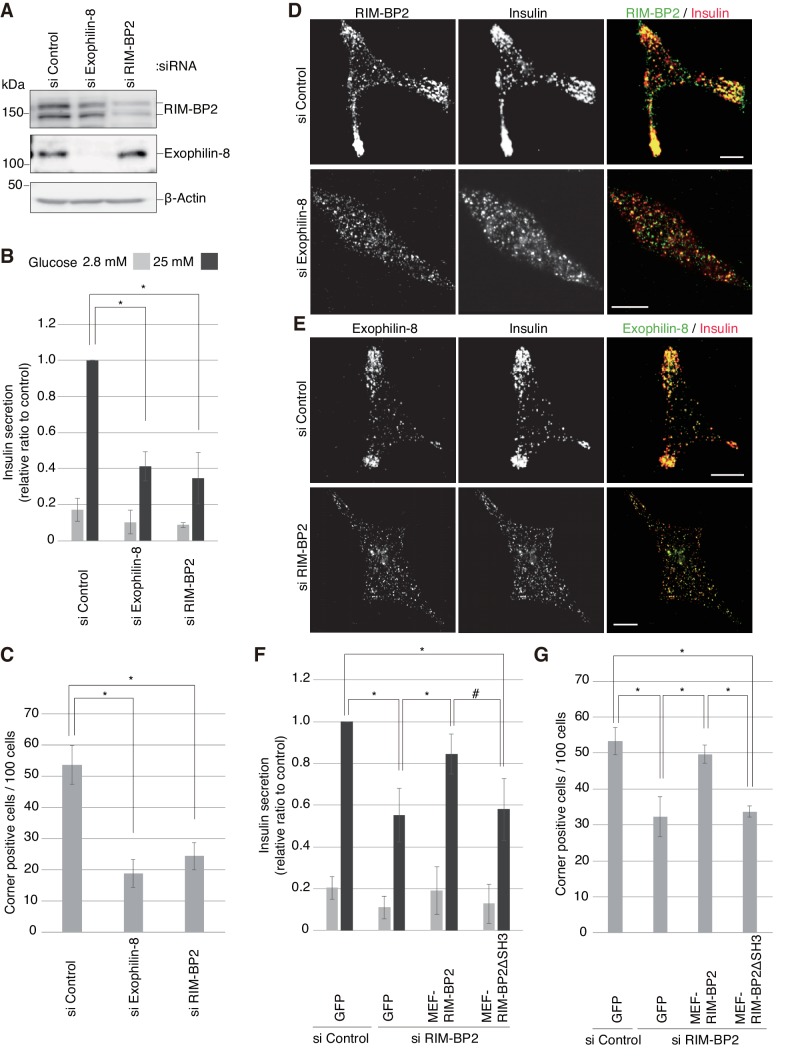
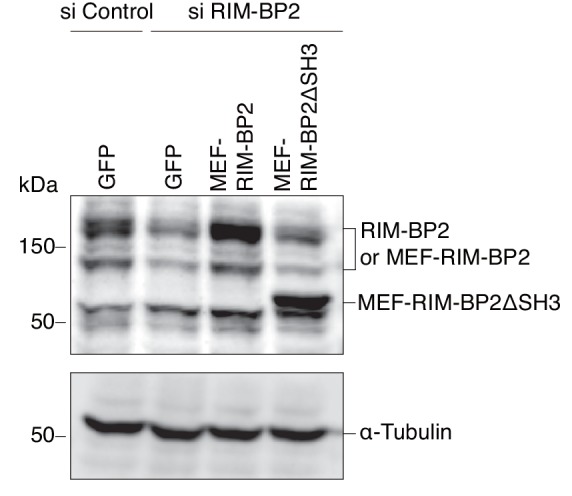
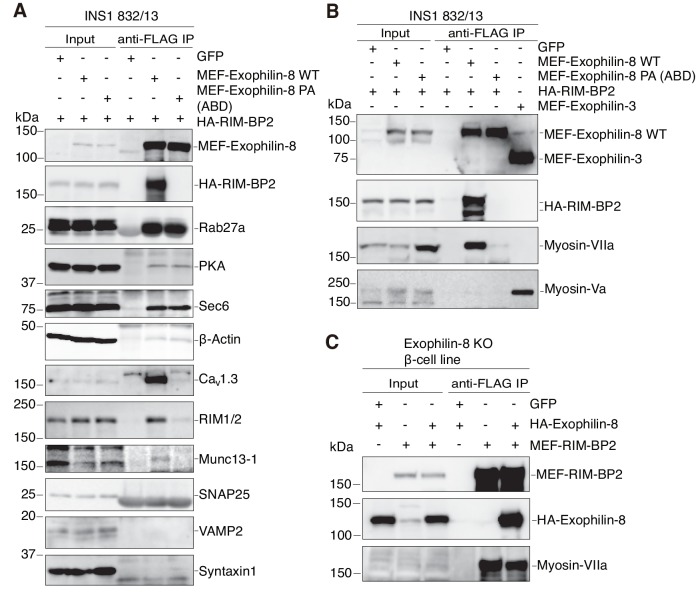
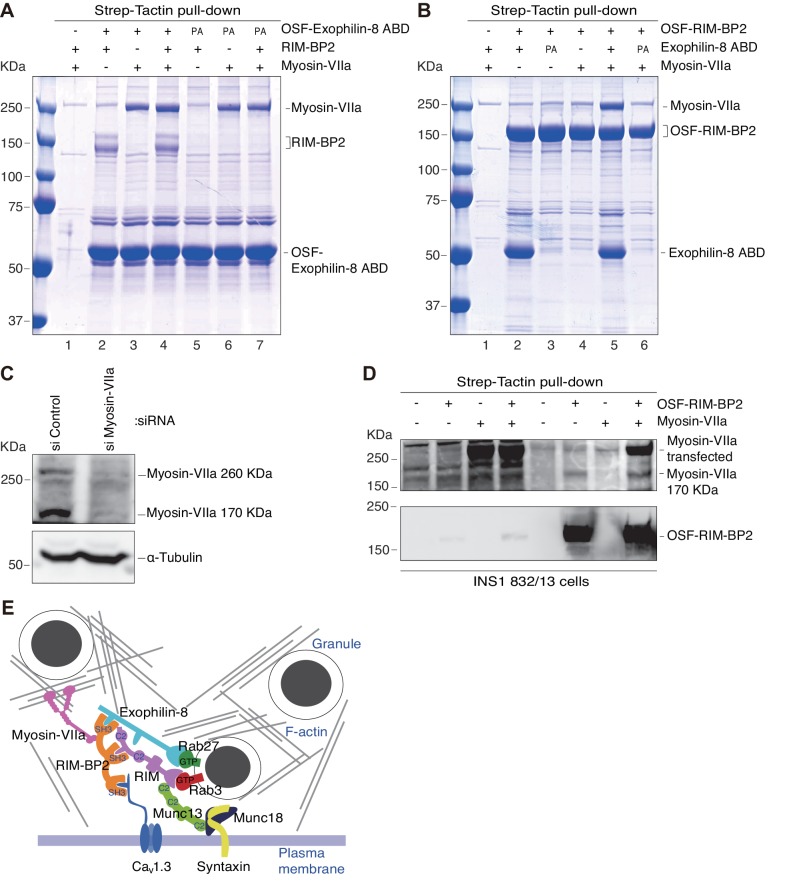
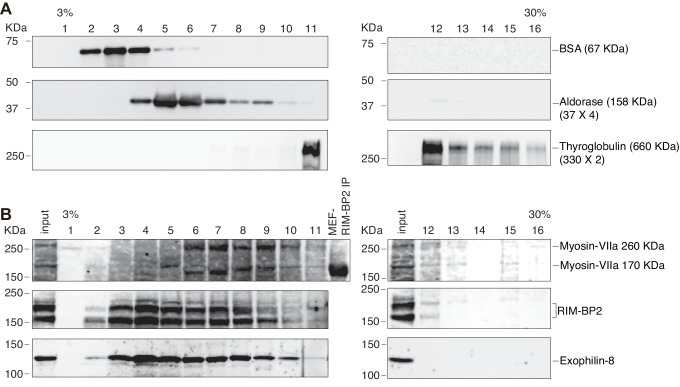
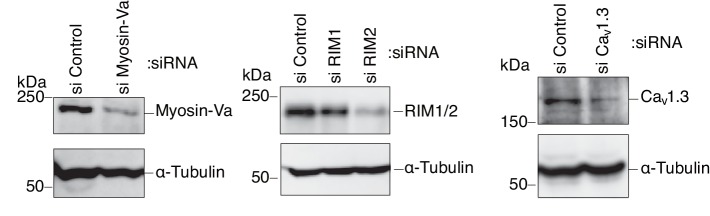
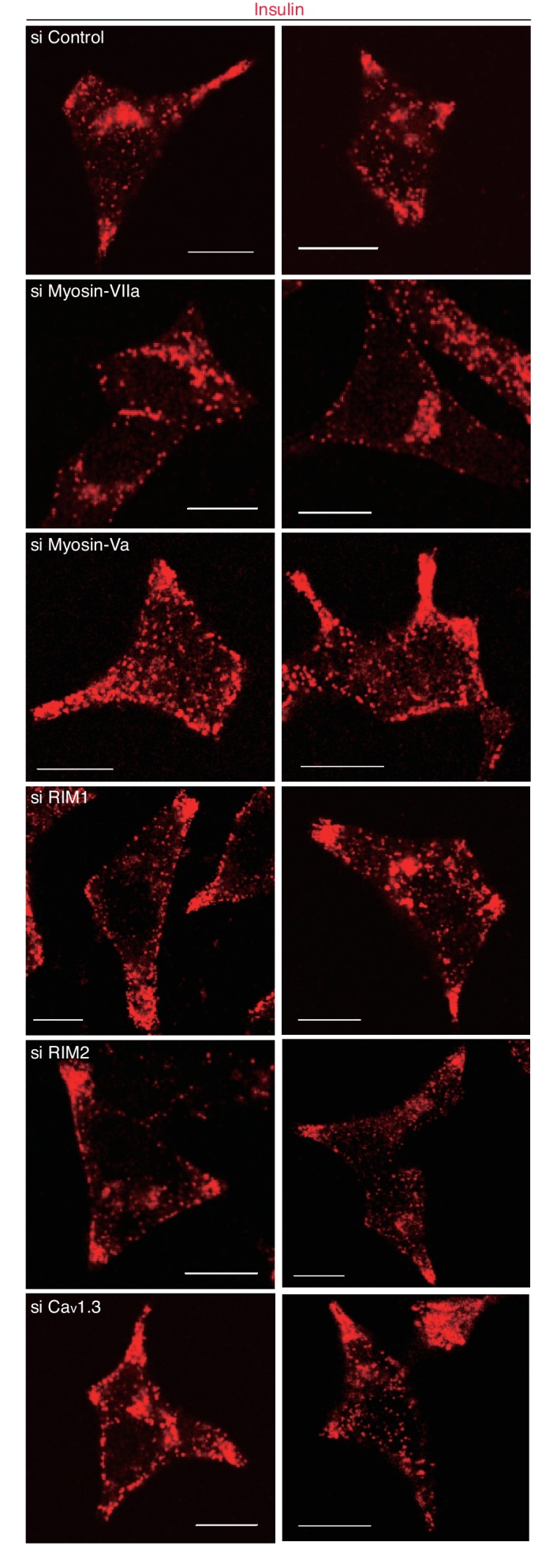
Exophilin-8 has been reported to play a role in anchoring secretory granules within the actin cortex, due to its direct binding activities to Rab27 on the granule membrane and to F-actin and its motor protein, myosin-Va. Here, we show that exophilin-8 accumulates granules in the cortical F-actin network not by direct interaction with myosin-Va, but by indirect interaction with a specific form of myosin-VIIa through its previously unknown binding partner, RIM-BP2. RIM-BP2 also associates with exocytic machinery, Ca1.3, RIM, and Munc13-1. Disruption of the exophilin-8-RIM-BP2-myosin-VIIa complex by ablation or knockdown of each component markedly decreases both the peripheral accumulation and exocytosis of granules. Furthermore, exophilin-8-null mouse pancreatic islets lose polarized granule localization at the β-cell periphery and exhibit impaired insulin secretion. This newly identified complex acts as a physical and functional scaffold and provides a mechanism supporting a releasable pool of granules within the F-actin network beneath the plasma membrane.
据报道,外嗜蛋白-8在将分泌颗粒锚定在肌动蛋白皮质中发挥作用,这是由于其与颗粒膜上的Rab27以及F-肌动蛋白及其运动蛋白肌球蛋白-Va具有直接结合活性。在此,我们表明,外嗜蛋白-8并非通过与肌球蛋白-Va直接相互作用,而是通过其先前未知的结合伴侣RIM-BP2与特定形式的肌球蛋白-VIIa间接相互作用,从而在皮质F-肌动蛋白网络中积累颗粒。RIM-BP2还与胞吐机制、Ca1.3、RIM和Munc13-1相关联。通过敲除或敲低每个组分来破坏外嗜蛋白-8-RIM-BP2-肌球蛋白-VIIa复合物,会显著降低颗粒的外周积累和胞吐作用。此外,外嗜蛋白-8基因敲除的小鼠胰岛在β细胞外周失去极化颗粒定位,并表现出胰岛素分泌受损。这种新鉴定的复合物作为一种物理和功能支架,提供了一种机制来支持质膜下方F-肌动蛋白网络内可释放颗粒池。