Chang Jessica T, Kumsta Caroline, Hellman Andrew B, Adams Linnea M, Hansen Malene
Program of Development, Aging and Regeneration, Sanford Burnham Prebys Medical Discovery Institute, La Jolla, United States.
Elife. 2017 Jul 4;6:e18459. doi: 10.7554/eLife.18459.
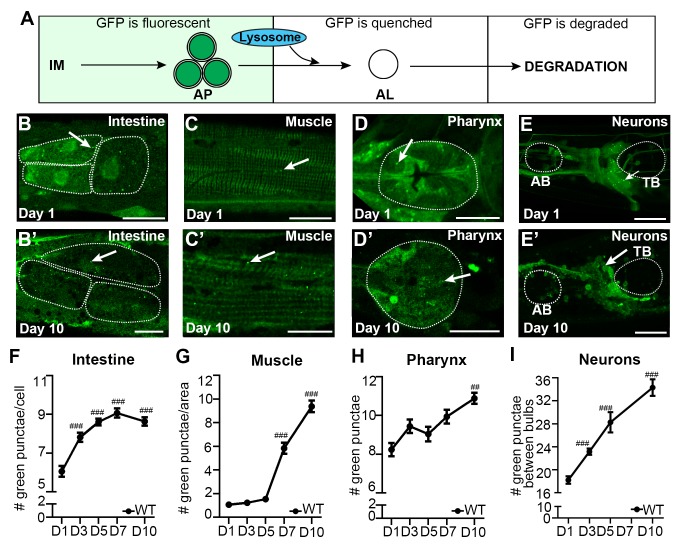
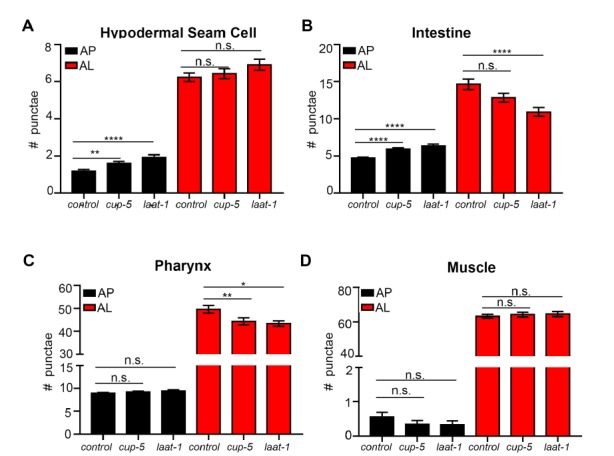
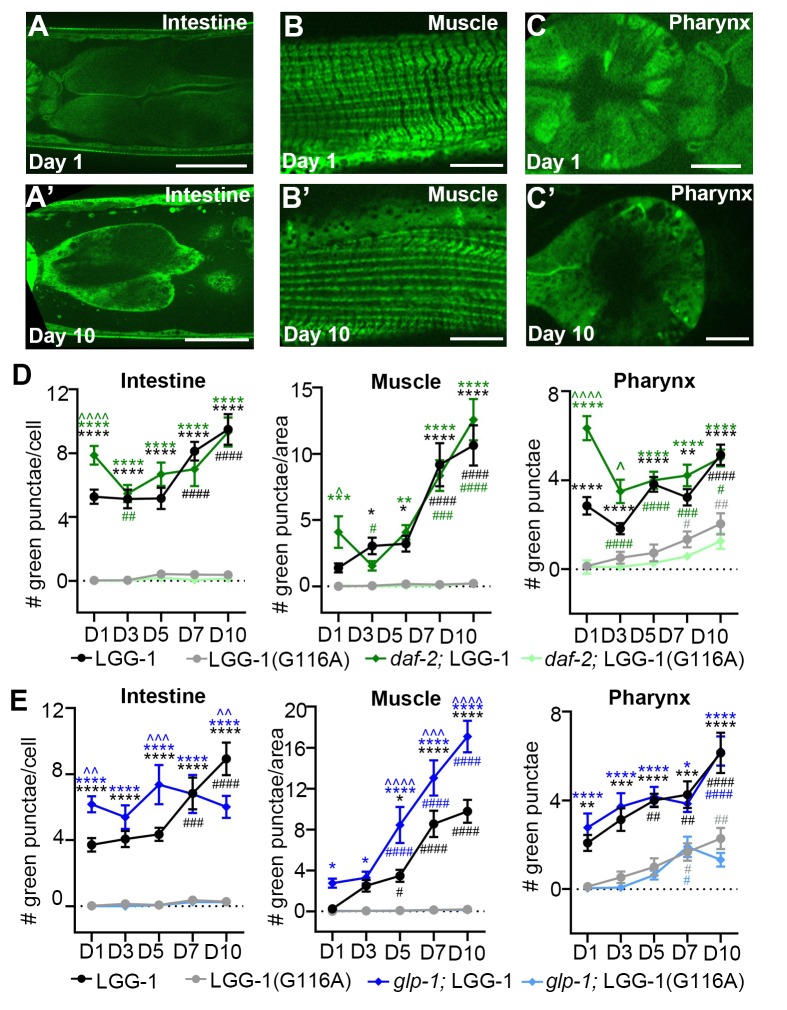
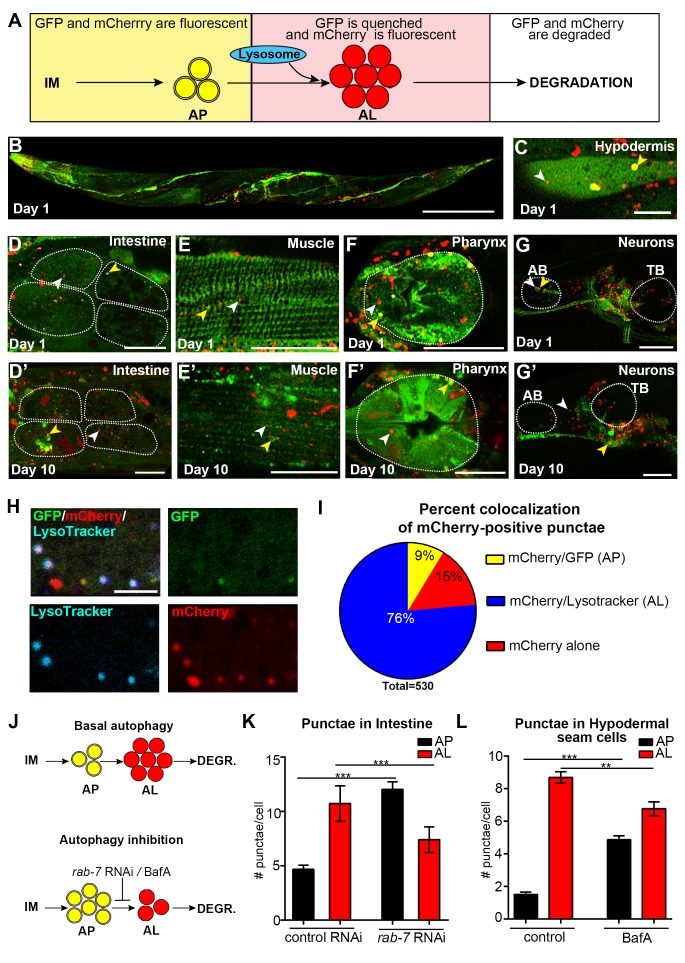
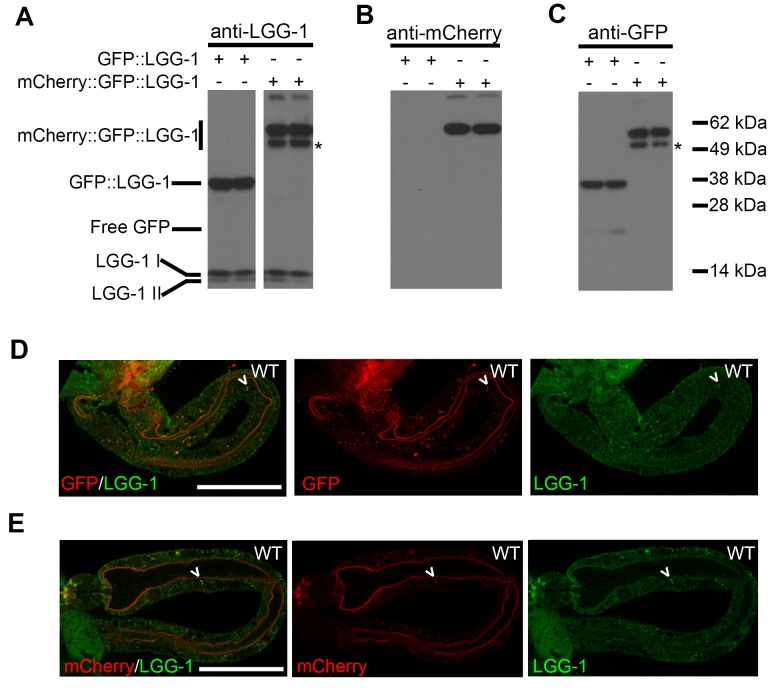
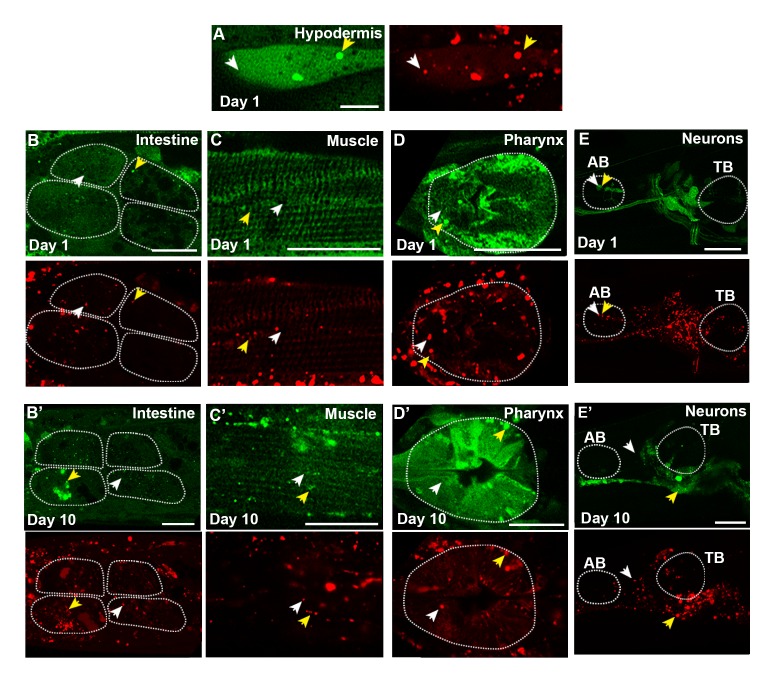
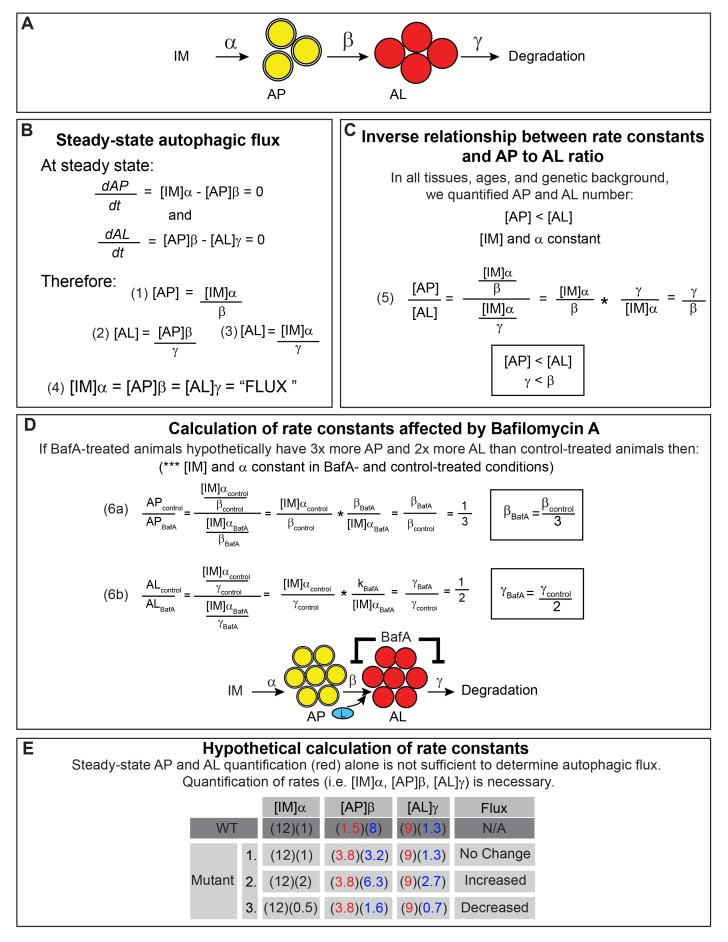
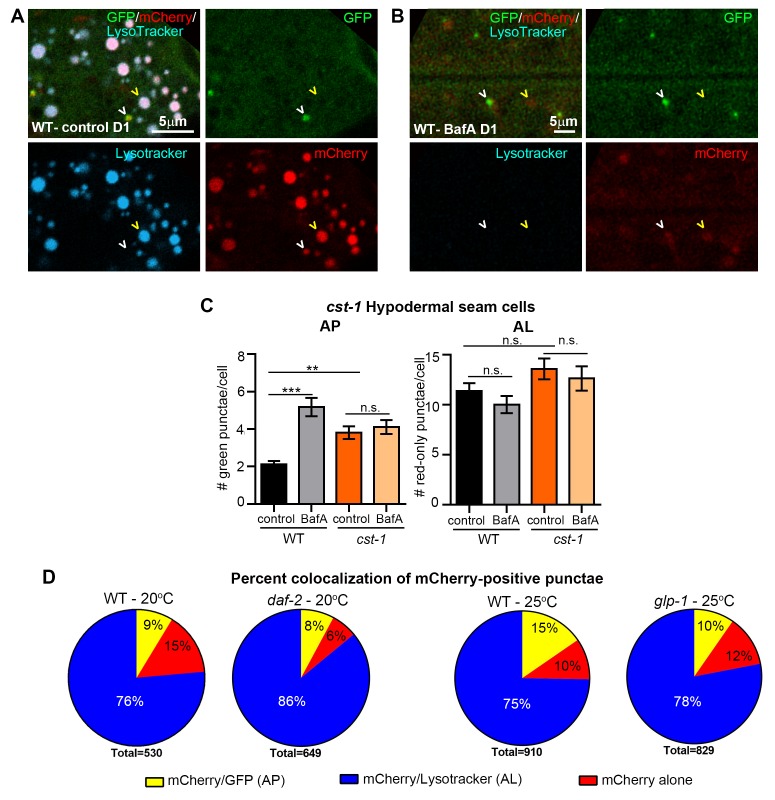
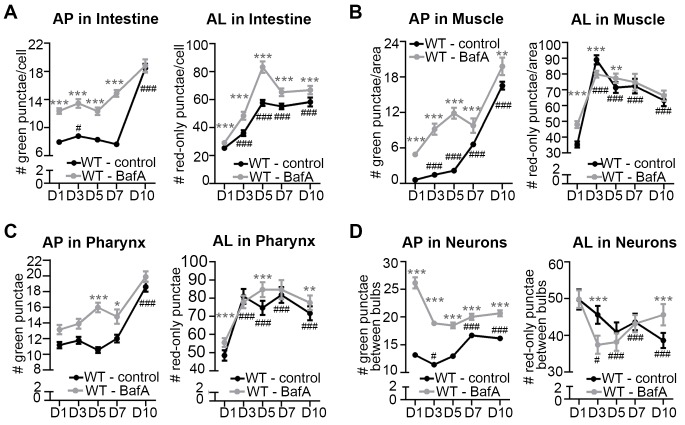
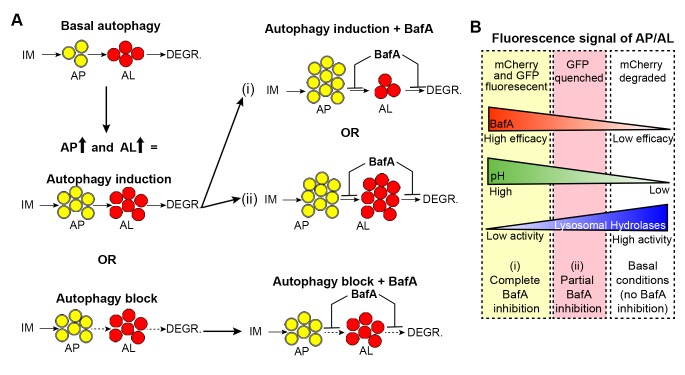
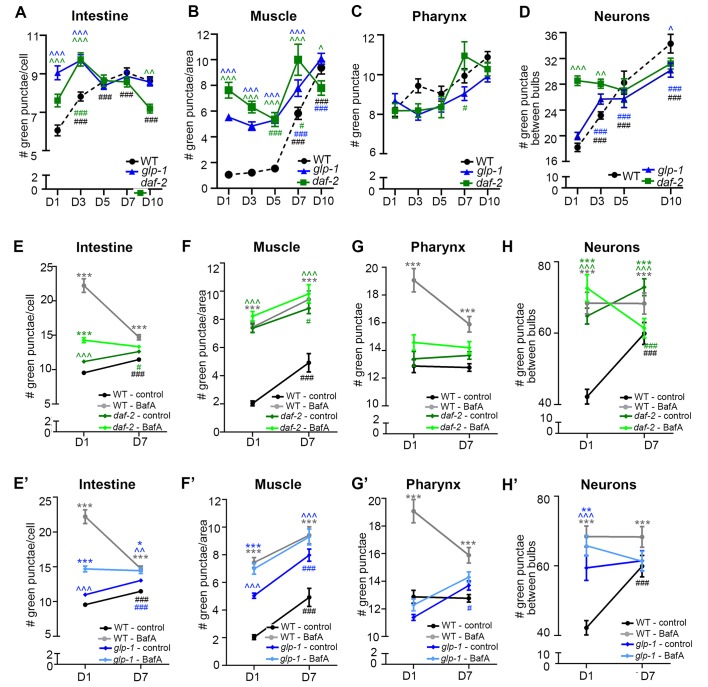
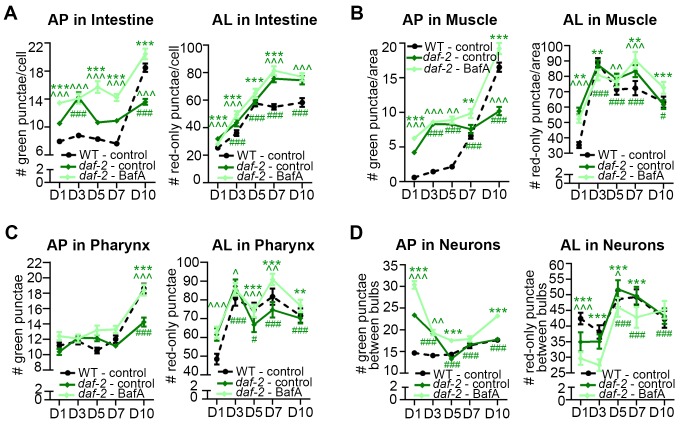
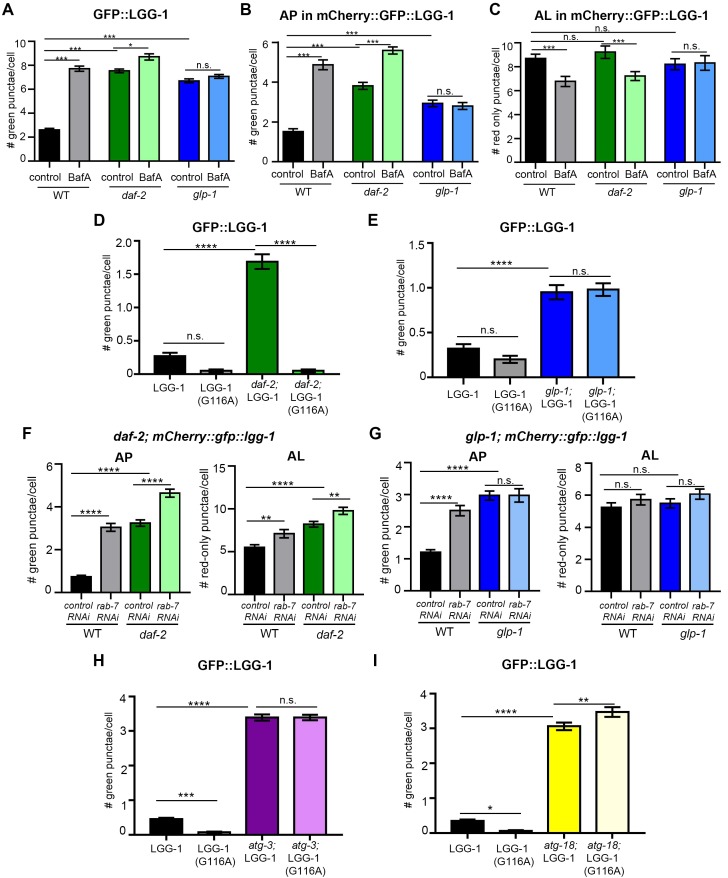
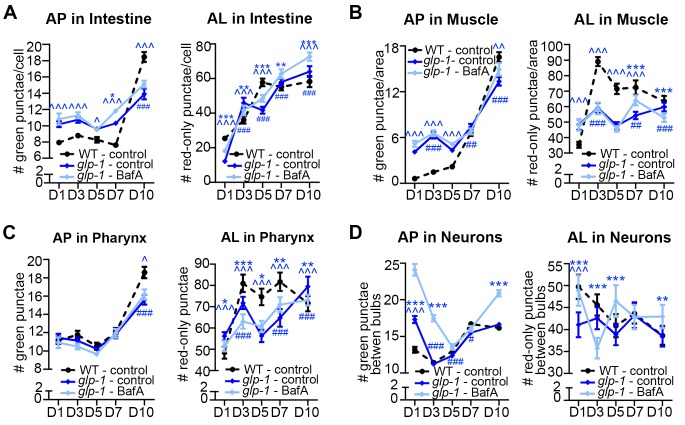
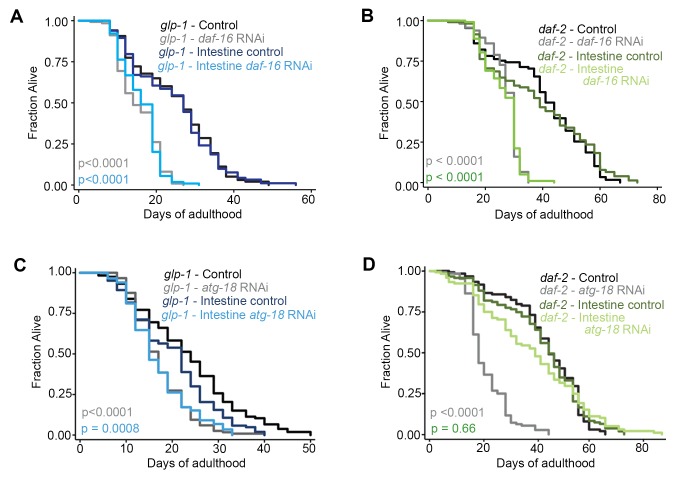
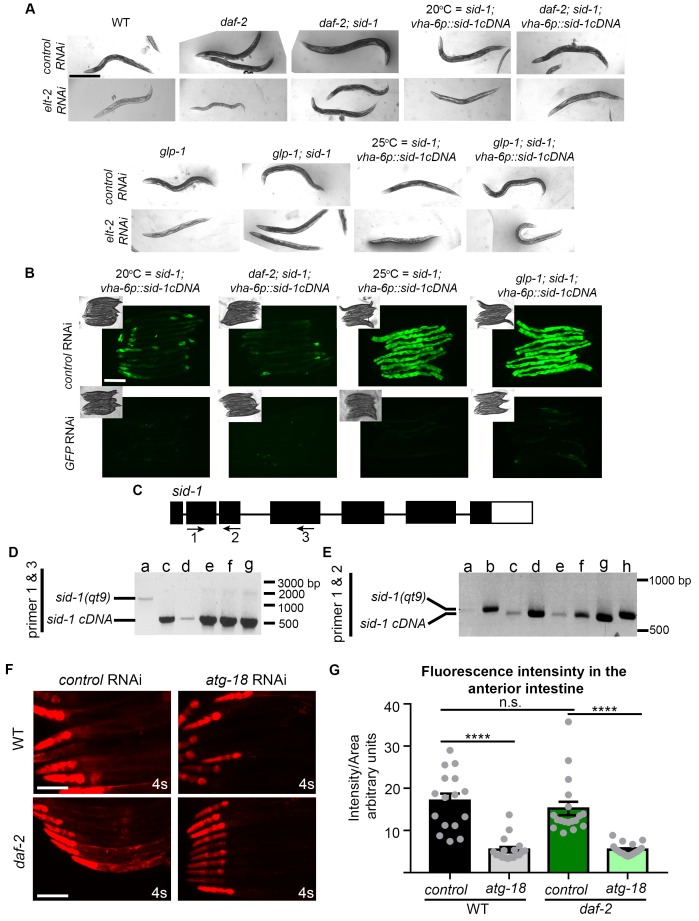
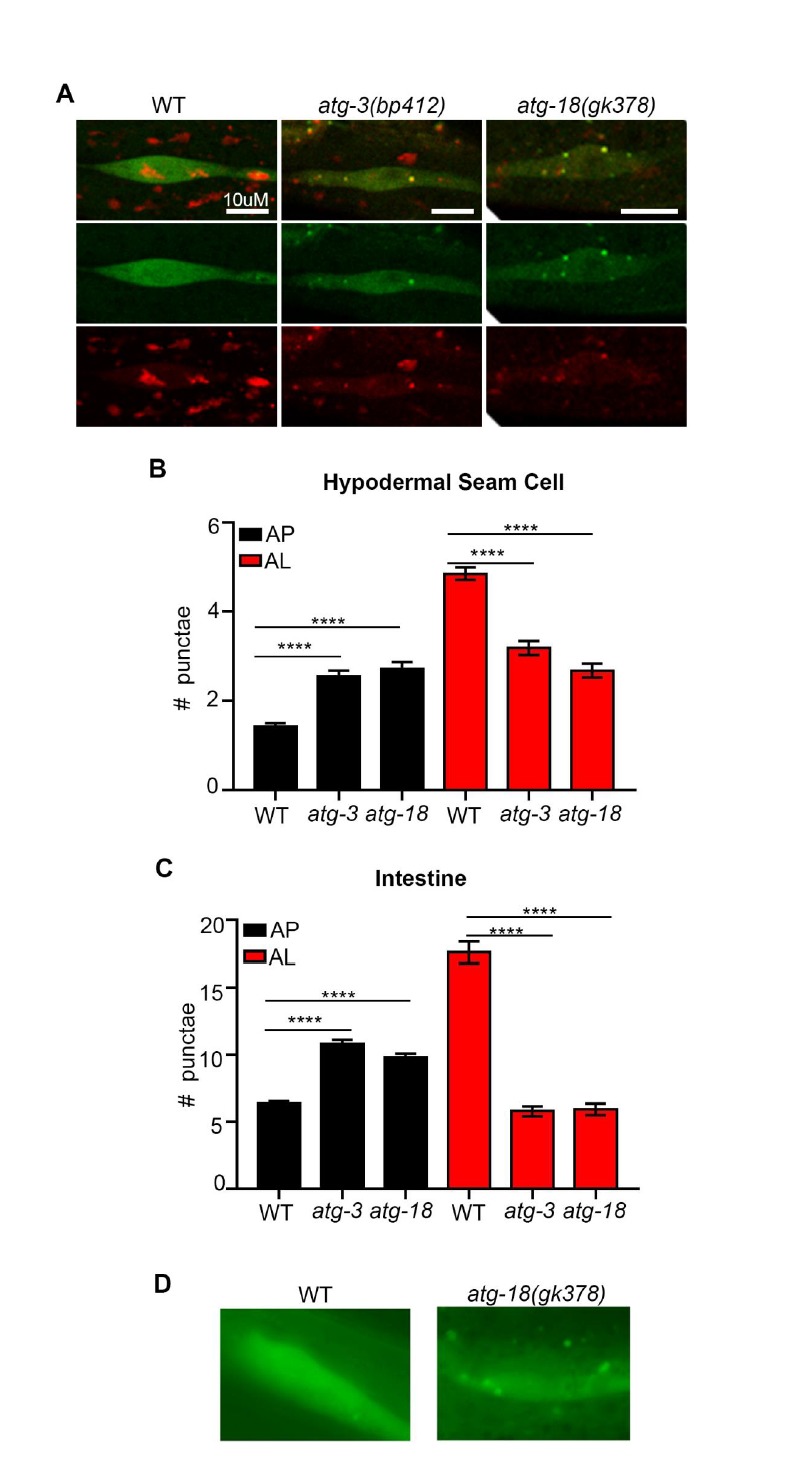
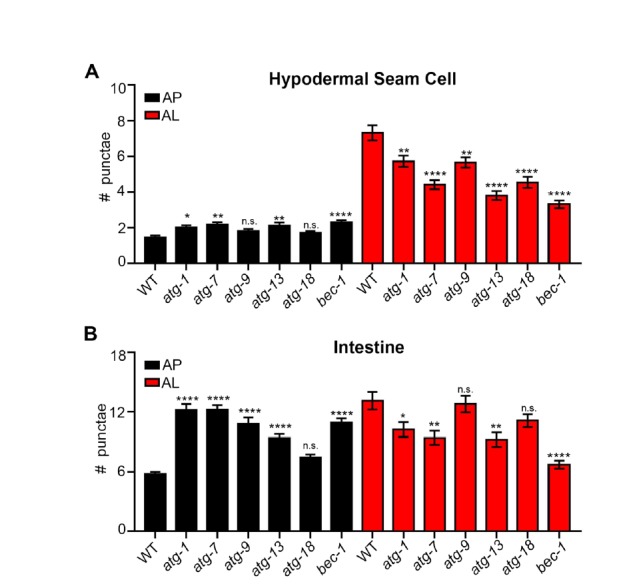
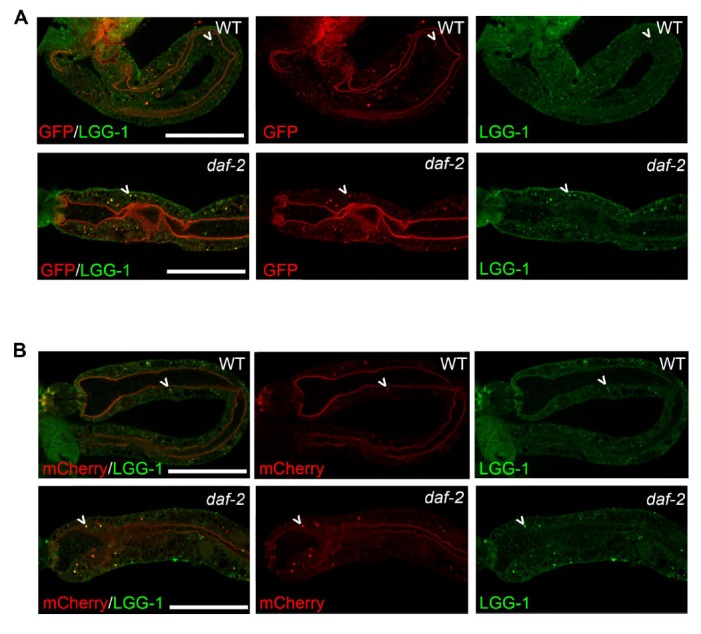
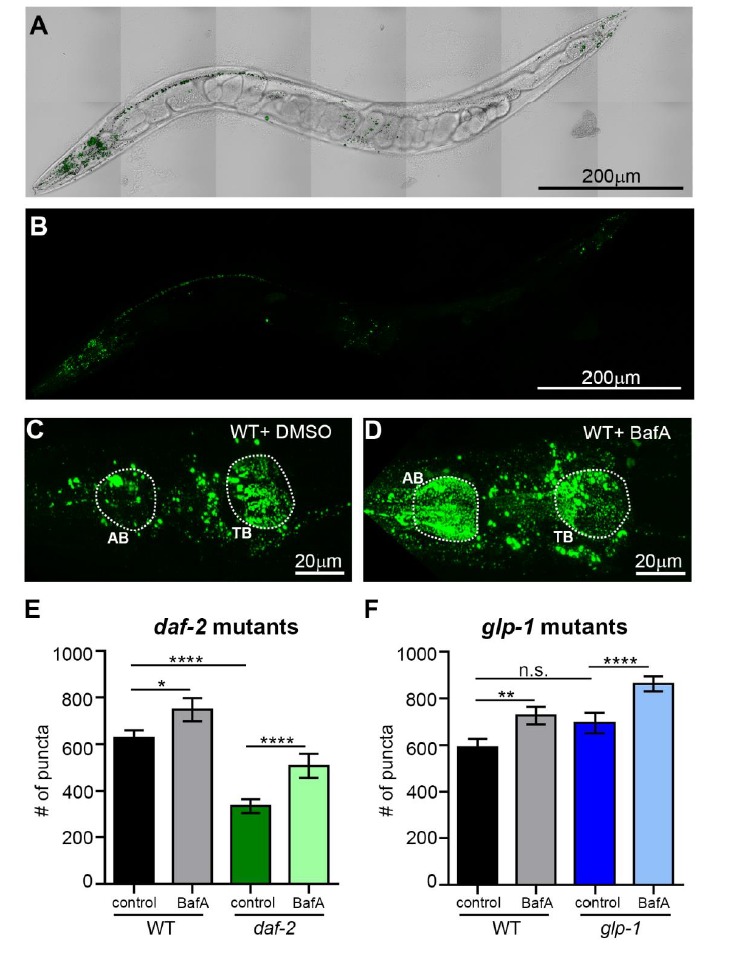
Autophagy has been linked to longevity in many species, but the underlying mechanisms are unclear. Using a GFP-tagged and a new tandem-tagged Atg8/LGG-1 reporter, we quantified autophagic vesicles and performed autophagic flux assays in multiple tissues of wild-type and long-lived /insulin/IGF-1 and /Notch mutants throughout adulthood. Our data are consistent with an age-related decline in autophagic activity in the intestine, body-wall muscle, pharynx, and neurons of wild-type animals. In contrast, and mutants displayed unique age- and tissue-specific changes in autophagic activity, indicating that the two longevity paradigms have distinct effects on autophagy during aging. Although autophagy appeared active in the intestine of both long-lived mutants, inhibition of intestinal autophagy significantly abrogated lifespan extension only in mutants. Collectively, our data suggest that autophagic activity normally decreases with age in whereas and long-lived mutants regulate autophagy in distinct spatiotemporal-specific manners to extend lifespan.
自噬与许多物种的寿命相关,但潜在机制尚不清楚。我们使用绿色荧光蛋白(GFP)标记的以及一种新的串联标记的Atg8/LGG-1报告基因,在成年期野生型以及长寿/胰岛素/胰岛素样生长因子-1(IGF-1)和Notch突变体的多个组织中对自噬小泡进行了定量,并进行了自噬通量分析。我们的数据与野生型动物肠道、体壁肌肉、咽部和神经元中自噬活性随年龄下降一致。相比之下,胰岛素/IGF-1和Notch突变体在自噬活性方面表现出独特的年龄和组织特异性变化,表明这两种长寿模式在衰老过程中对自噬有不同影响。尽管在两种长寿突变体的肠道中自噬似乎都很活跃,但仅在胰岛素/IGF-1突变体中,抑制肠道自噬会显著消除寿命延长。总体而言,我们的数据表明,在野生型中自噬活性通常随年龄降低,而胰岛素/IGF-1和Notch长寿突变体以不同的时空特异性方式调节自噬以延长寿命。