Shute Elen, Prideaux Gavin J, Worthy Trevor H
School of Biological Sciences, Flinders University, GPO Box 2100, Adelaide, South Australia 5001, Australia.
R Soc Open Sci. 2017 Jun 14;4(6):170233. doi: 10.1098/rsos.170233. eCollection 2017 Jun.
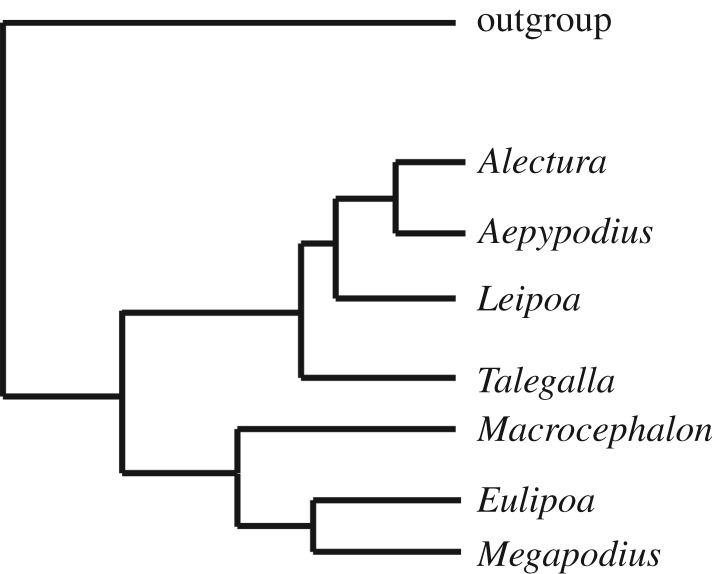
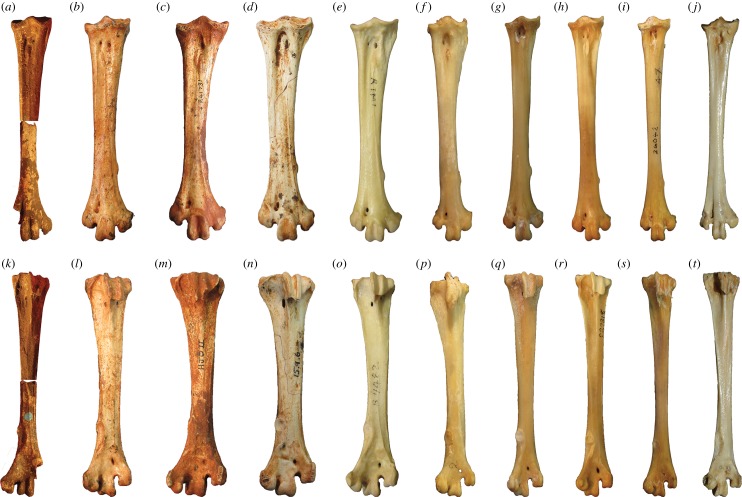
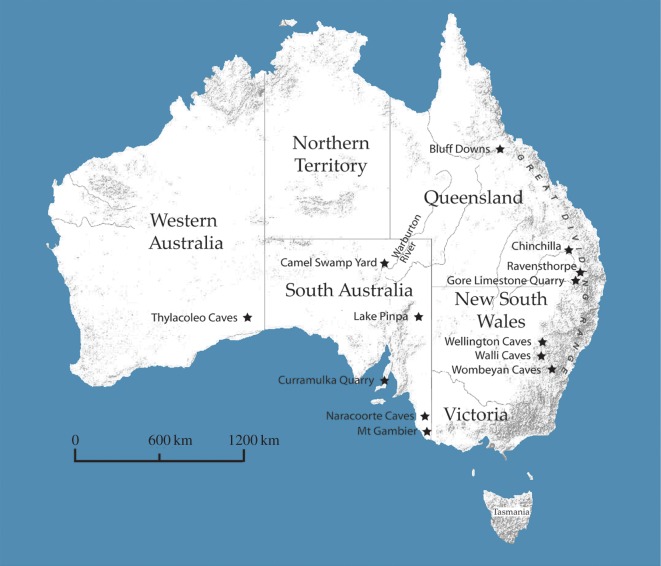
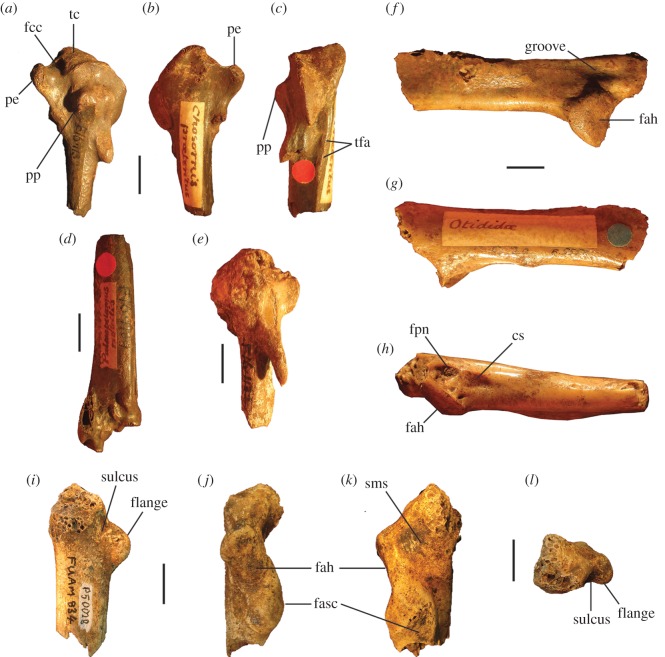
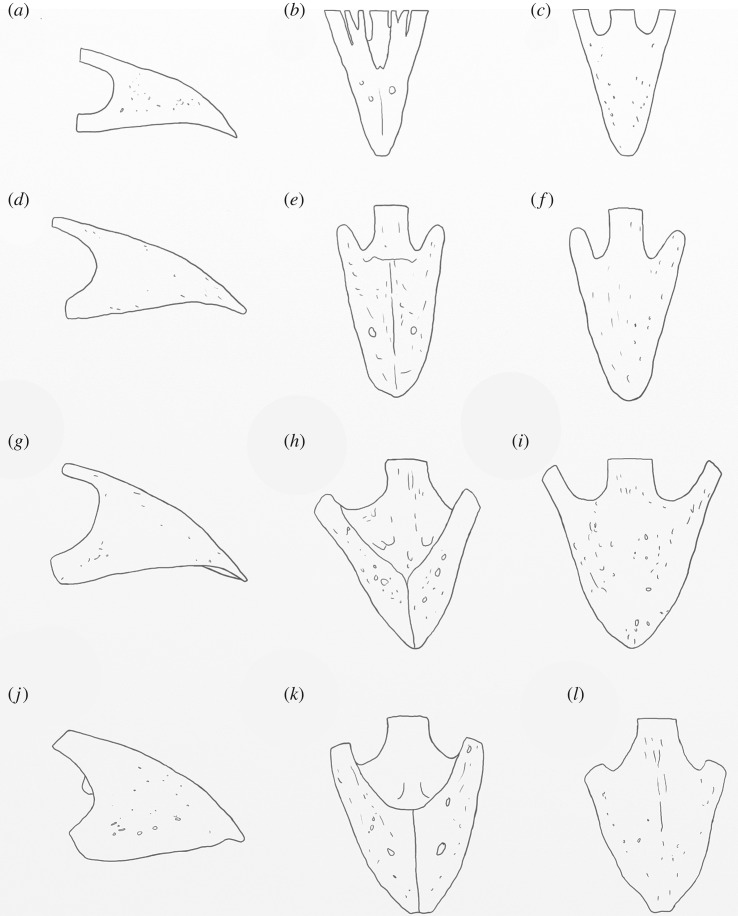
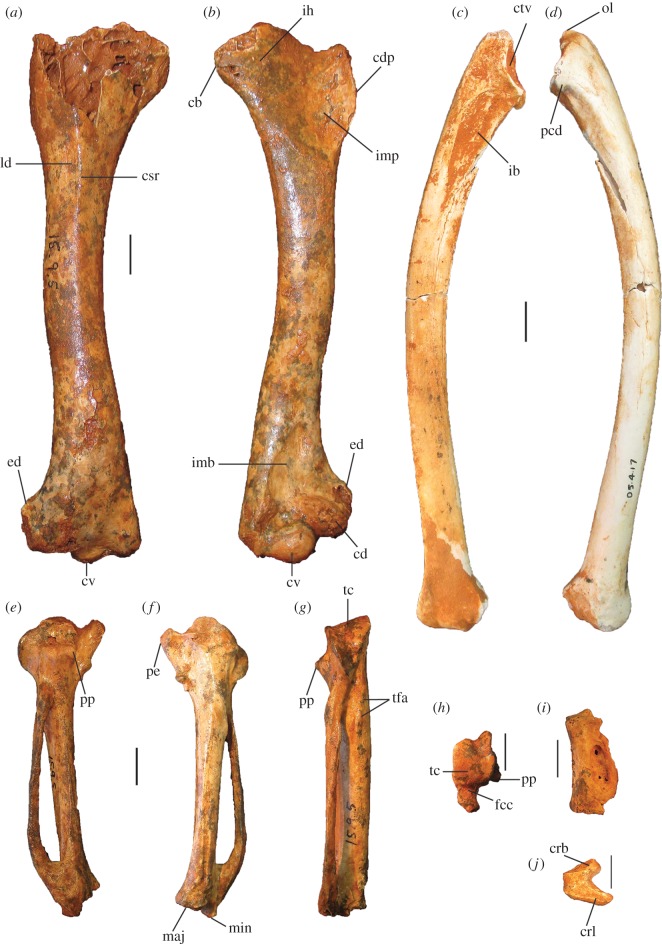
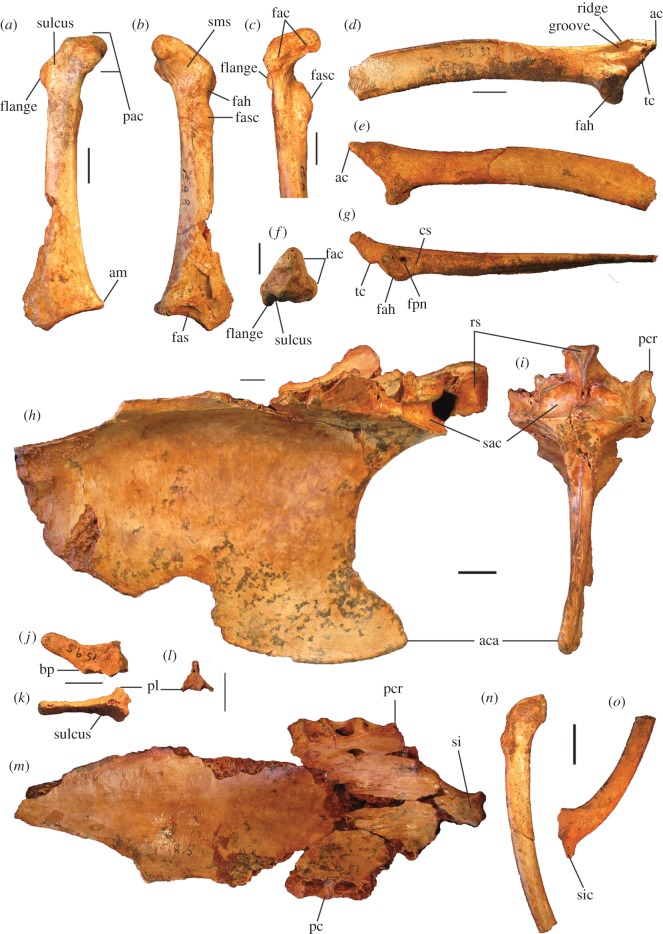
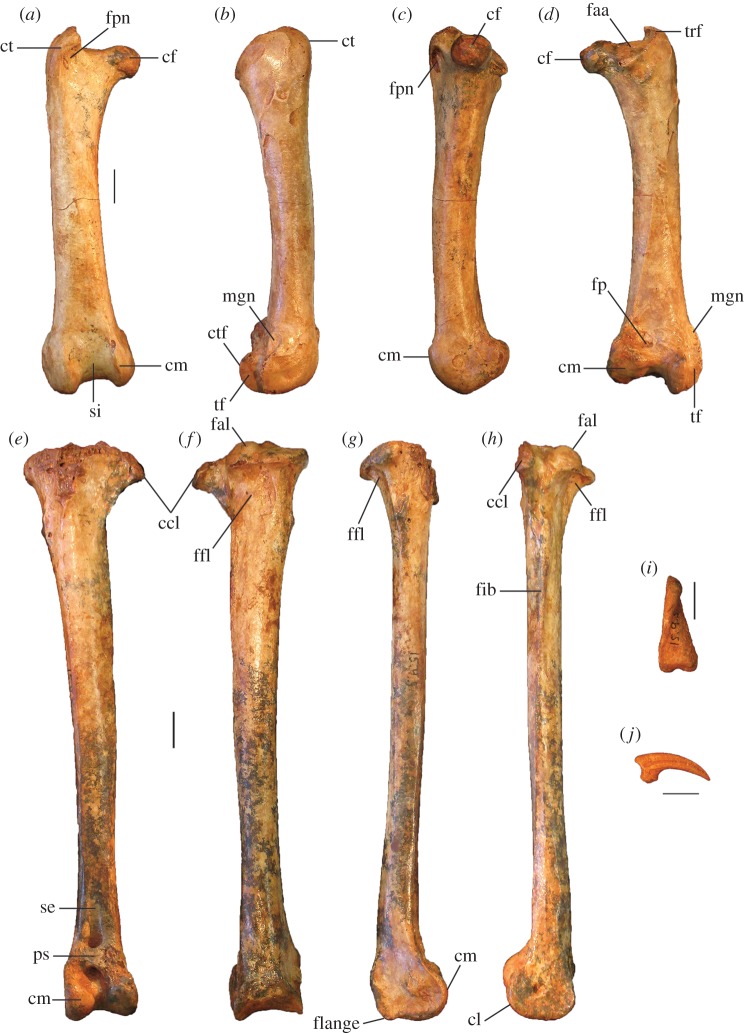
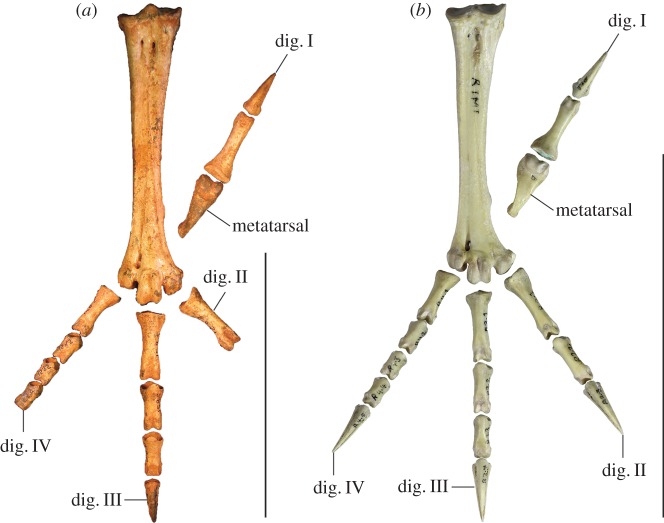
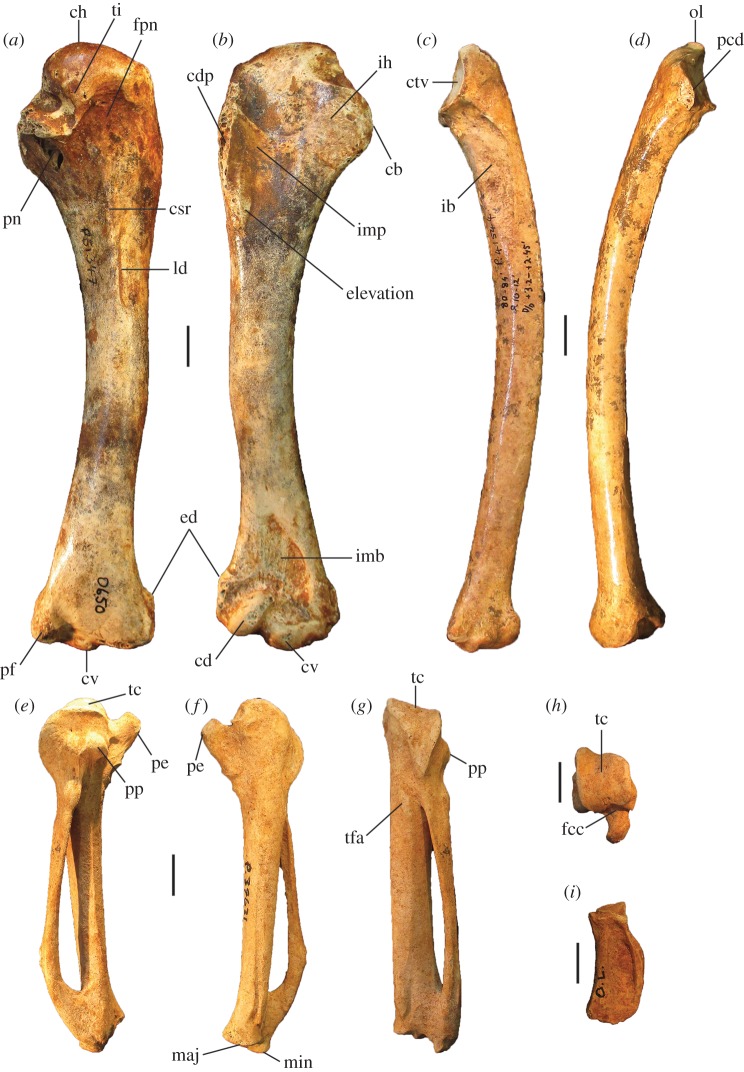
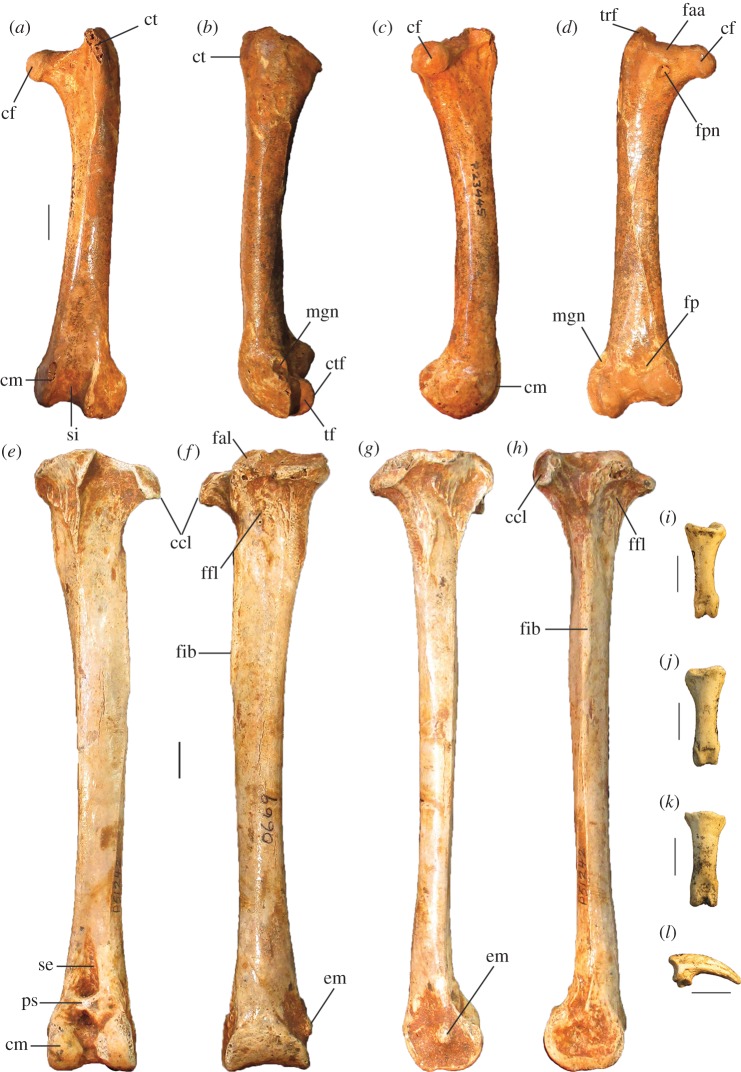
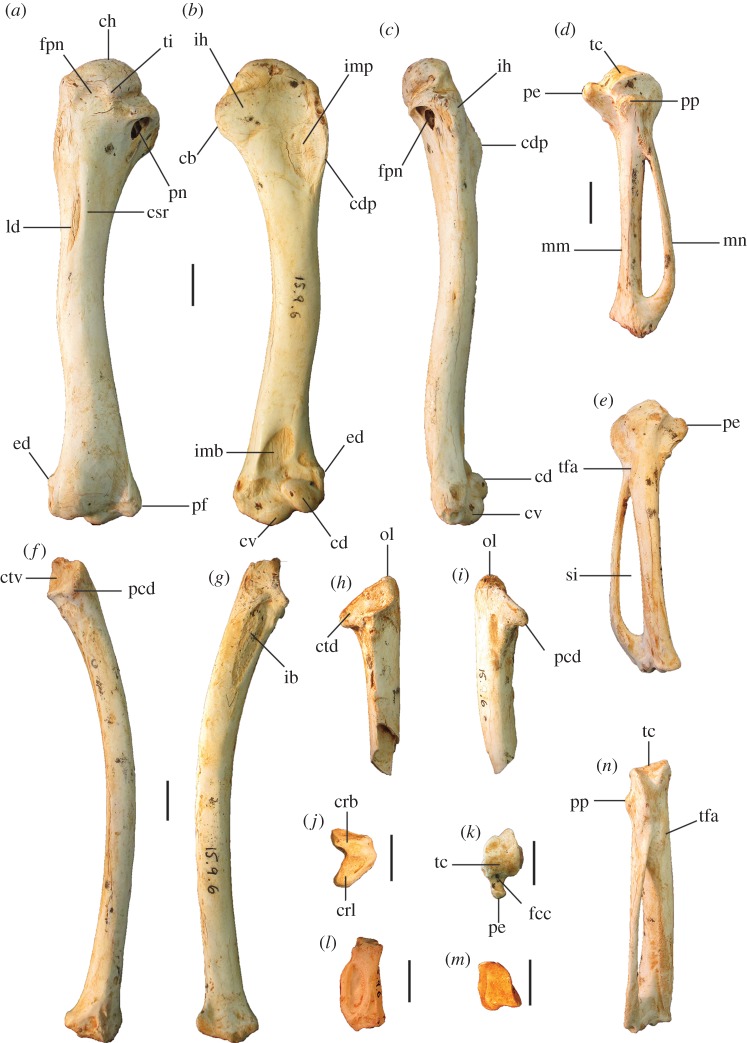
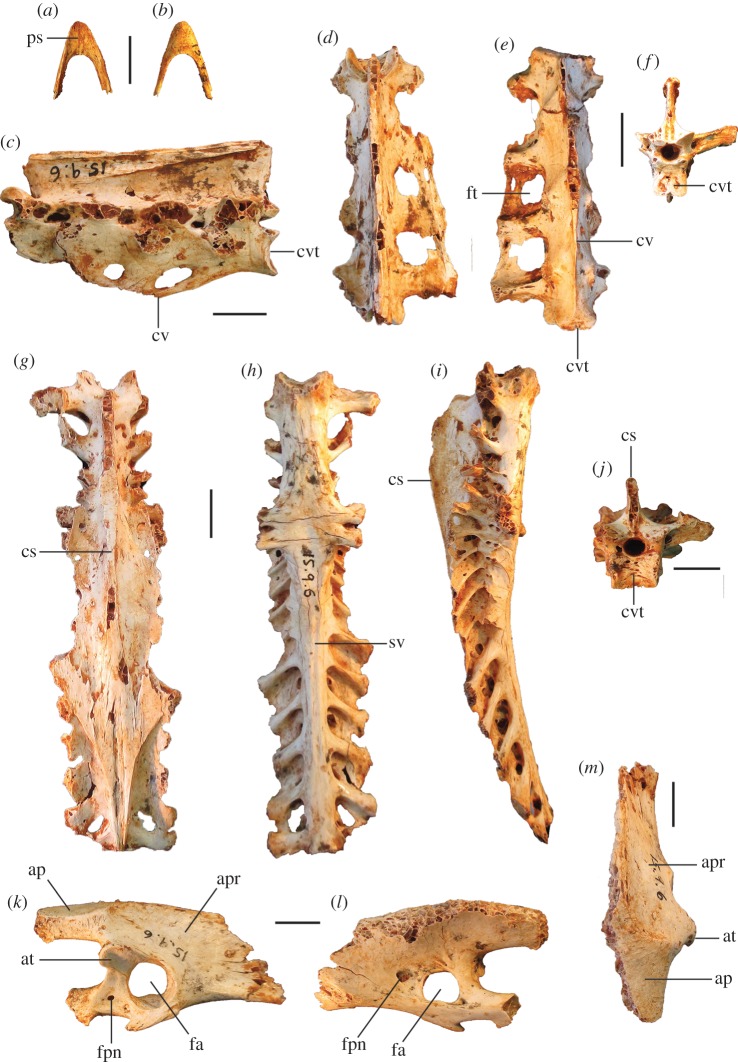
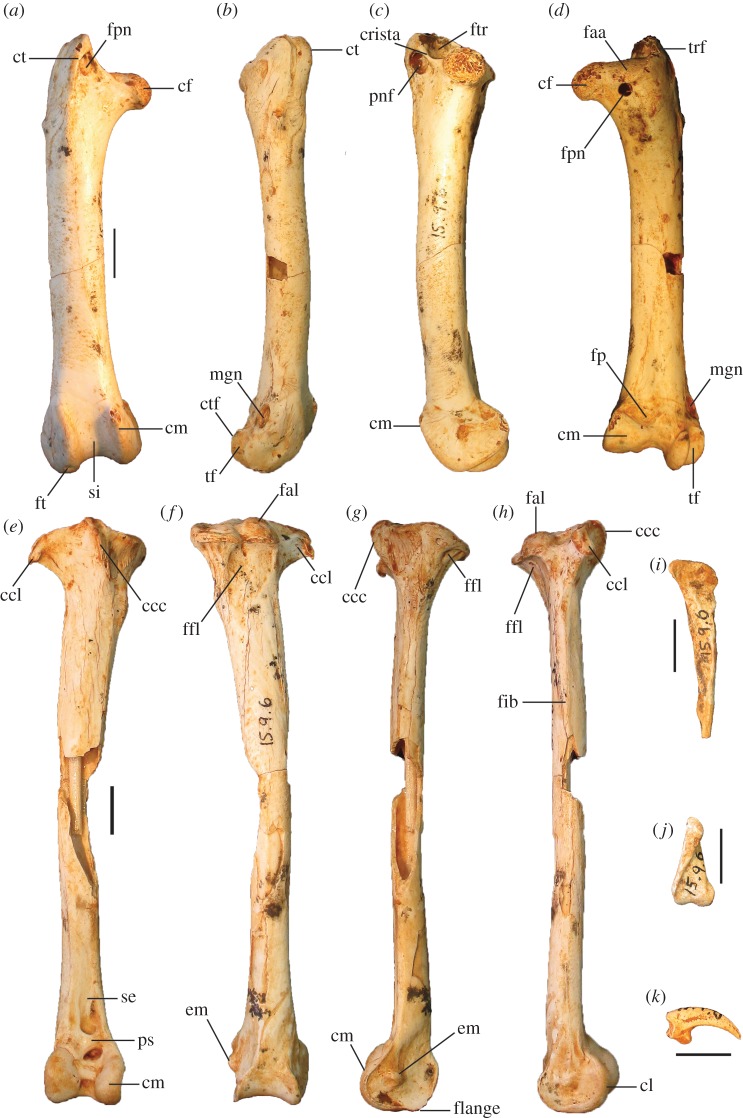
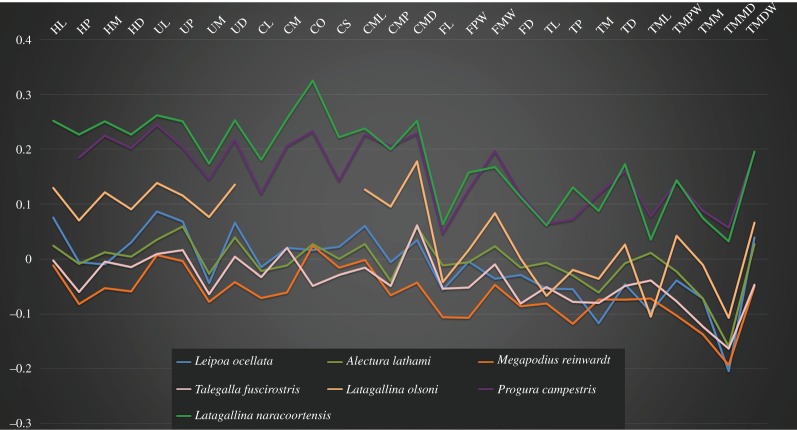
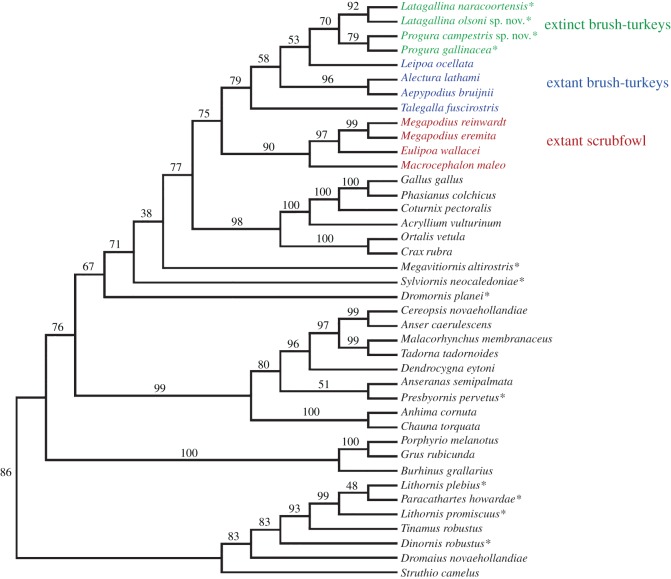
Megapodes are unusual galliform birds that use passive heat sources to incubate their eggs. Evolutionary relationships of extant megapode taxa have become clearer with the advent of molecular analyses, but the systematics of large, extinct forms (, ) from the late Cenozoic of Australia has been a source of confusion. It was recently suggested that the two species of were synonymous, and that this taxon dwarfed into the extant malleefowl in the Late Pleistocene. Here, we review previously described fossils along with newly discovered material from several localities, and present a substantial taxonomic revision. We show that and are generically distinct, describe two new species of megapode from the Thylacoleo Caves of south-central Australia, and a new genus from Curramulka Quarry in southern Australia. We also show that was contemporaneous with larger species. Our phylogenetic analysis places four extinct taxa in a derived clade with the extant Australo-Papuan brush-turkeys , , and . Therefore, diversity of brush-turkeys halved during the Quaternary, matching extinction rates of scrubfowl in the Pacific. Unlike extant brush-turkeys, all the extinct taxa appear to have been burrow-nesters.
冢雉是一种不同寻常的鸡形目鸟类,它们利用被动热源来孵化卵。随着分子分析的出现,现存冢雉类群的进化关系变得更加清晰,但来自澳大利亚晚新生代的大型已灭绝种类( )的系统分类一直是个令人困惑的问题。最近有人提出, 的两个物种是同义词,并且这个分类单元在晚更新世时演变成了现存的眼斑冢雉。在这里,我们回顾了之前描述的化石以及来自几个地点的新发现材料,并进行了一次重大的分类修订。我们表明, 和 在属级上是不同的,描述了来自澳大利亚中南部袋狮洞穴的两种新的冢雉物种,以及来自澳大利亚南部库拉穆尔卡采石场的一个新属。我们还表明, 与更大的物种是同时期的。我们的系统发育分析将四个已灭绝的分类单元置于一个衍生分支中,与现存的澳大利亚 - 巴布亚灌丛火鸡 、 、 和 在一起。因此,灌丛火鸡的多样性在第四纪减半,与太平洋地区 scrubfowl 的灭绝率相当。与现存的灌丛火鸡不同,所有已灭绝的分类单元似乎都是穴巢筑巢者。