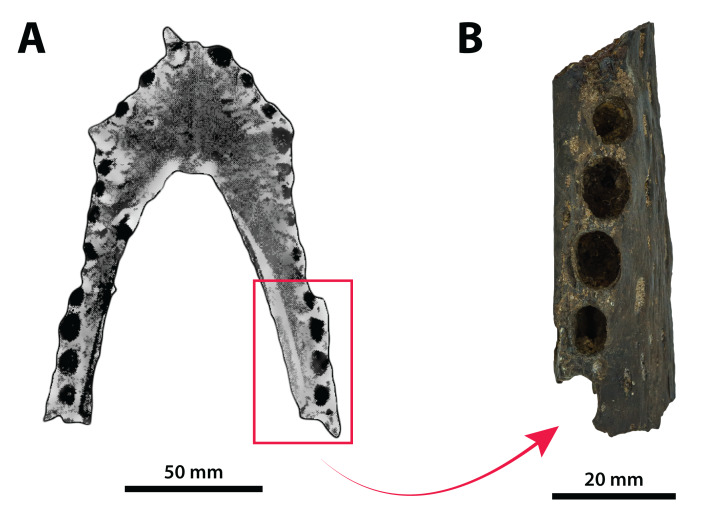
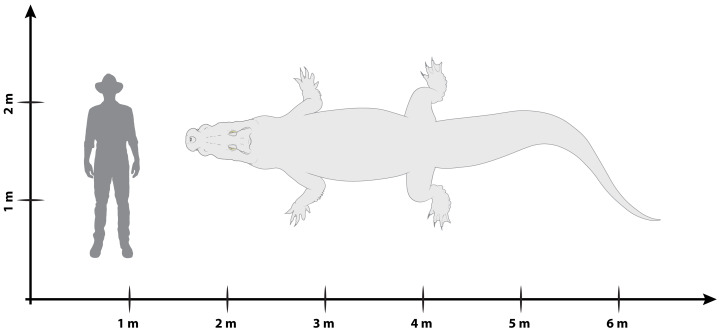
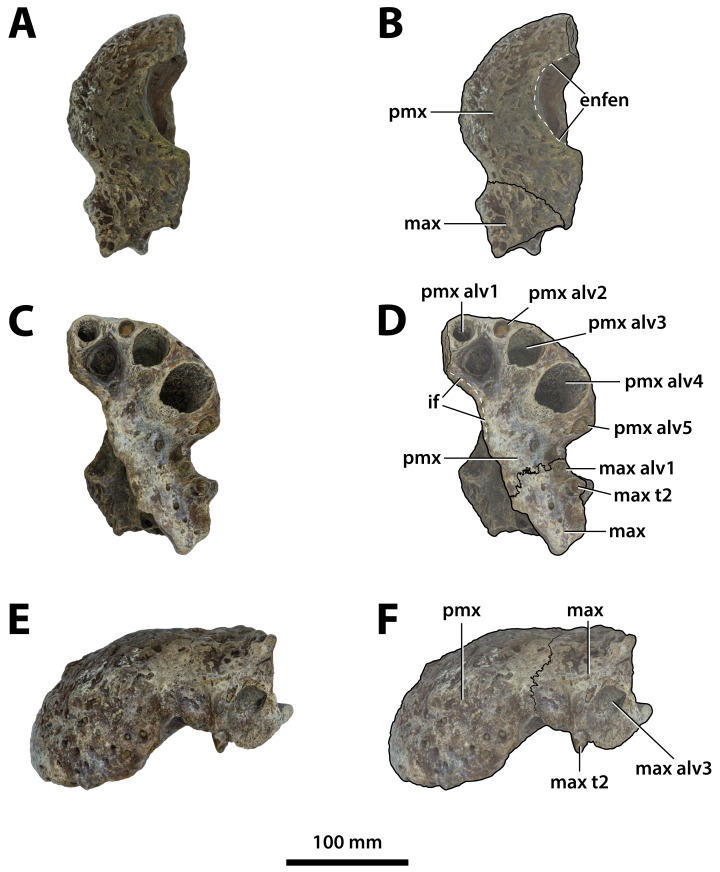
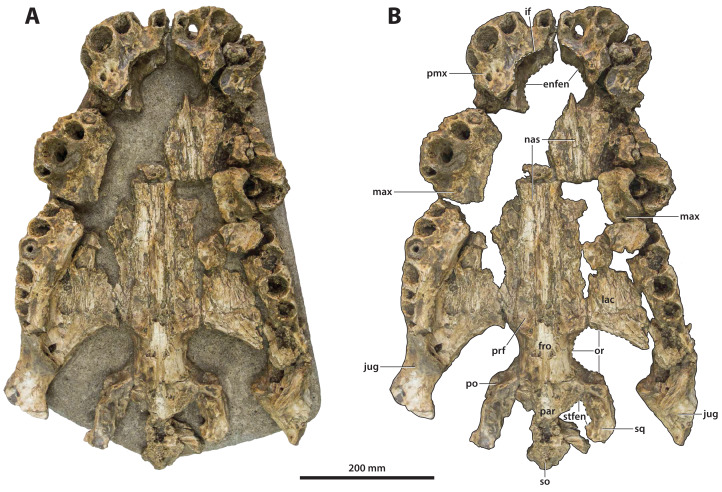
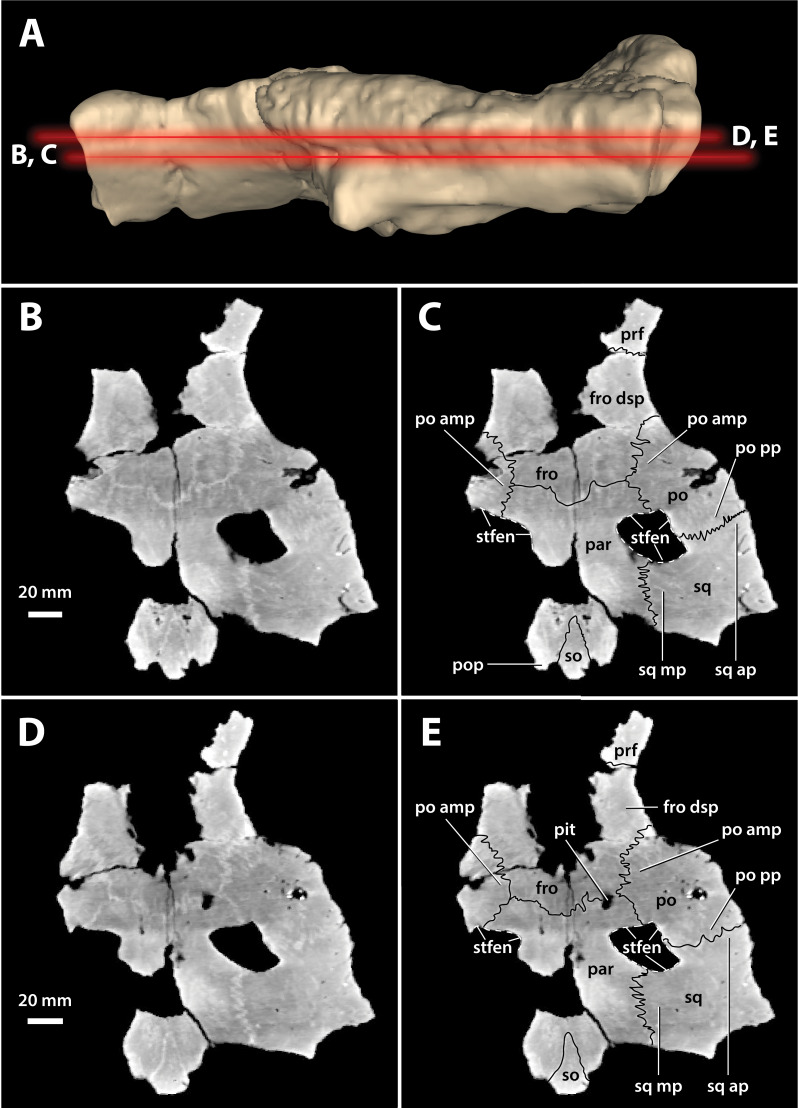
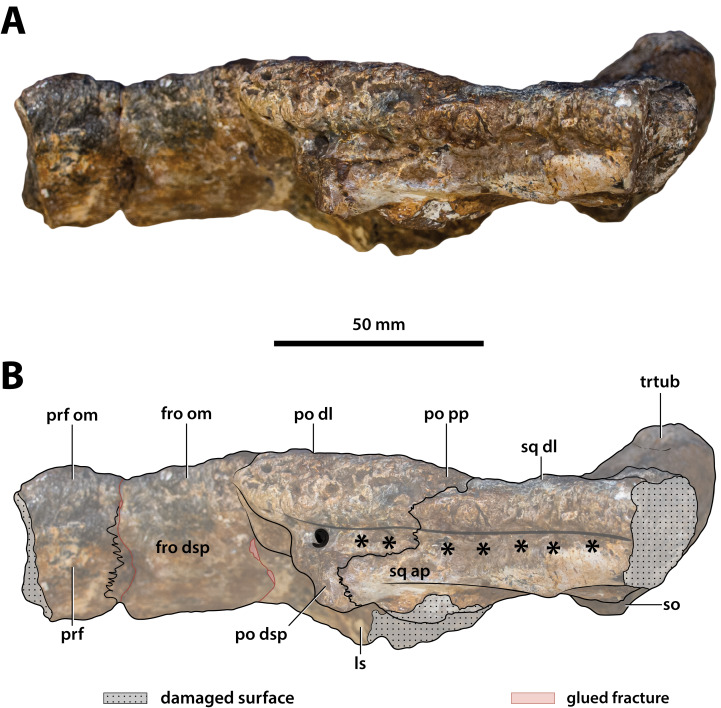
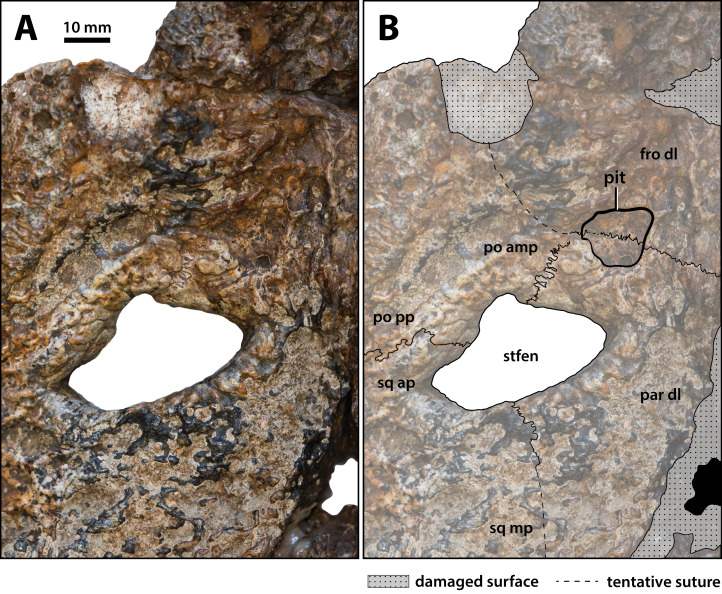
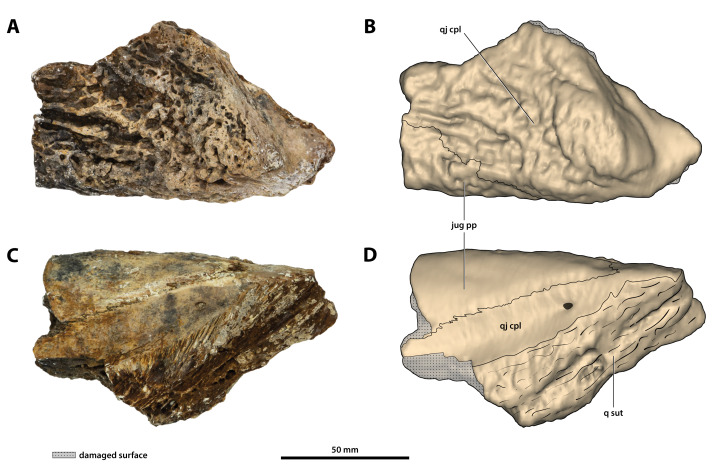
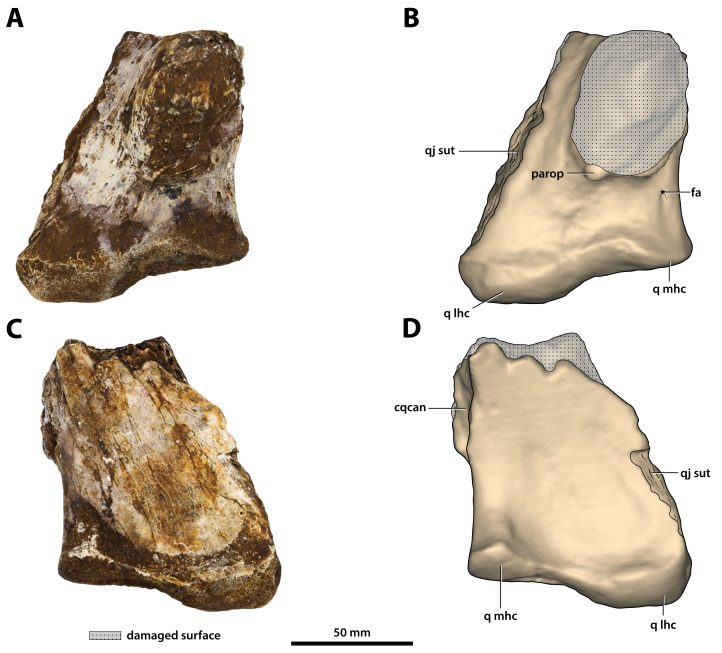
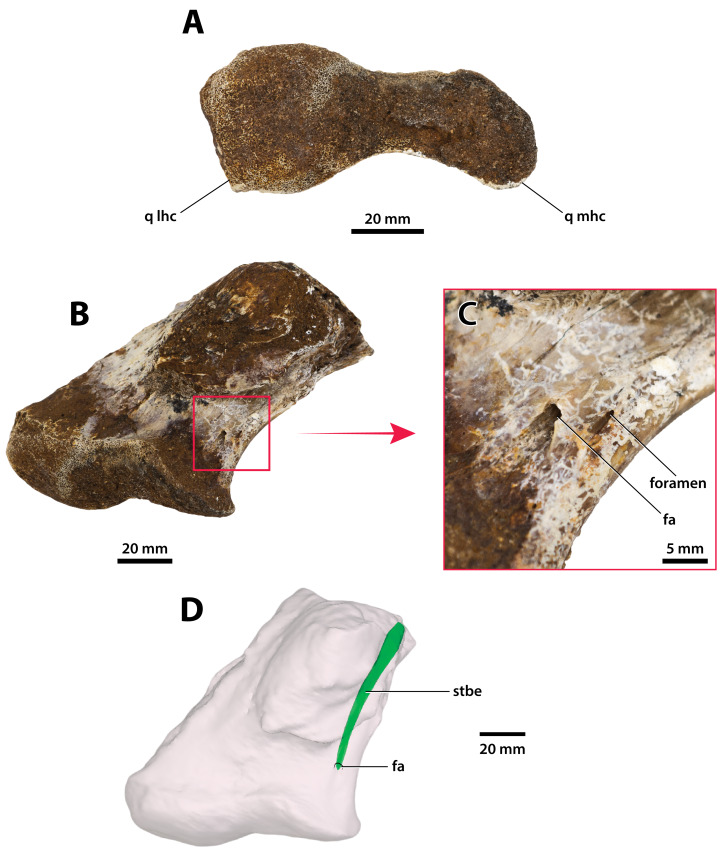
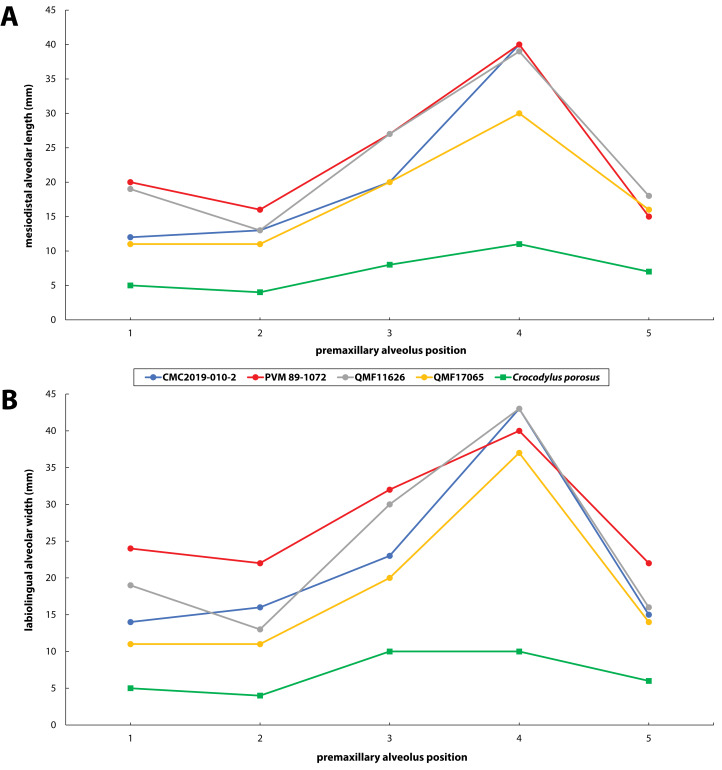
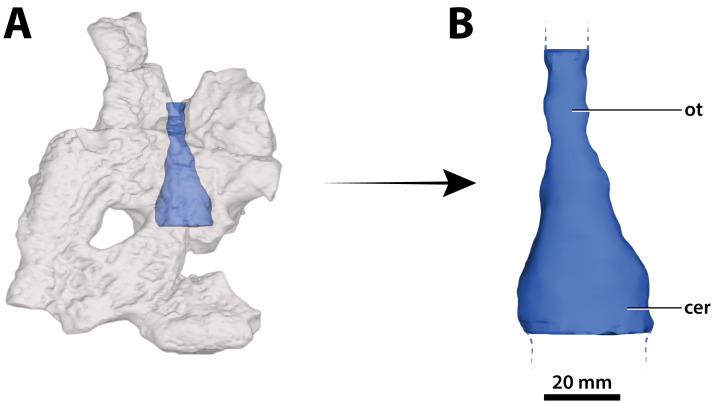
Ristevski Jorgo, Yates Adam M, Price Gilbert J, Molnar Ralph E, Weisbecker Vera, Salisbury Steven W
School of Biological Sciences, The Univeristy of Queensland, Brisbane, QLD, Australia.
Museum of Central Australia, Museum and Art Gallery of the Northern Territory, Alice Springs, NT, Australia.
PeerJ. 2020 Dec 21;8:e10466. doi: 10.7717/peerj.10466. eCollection 2020.
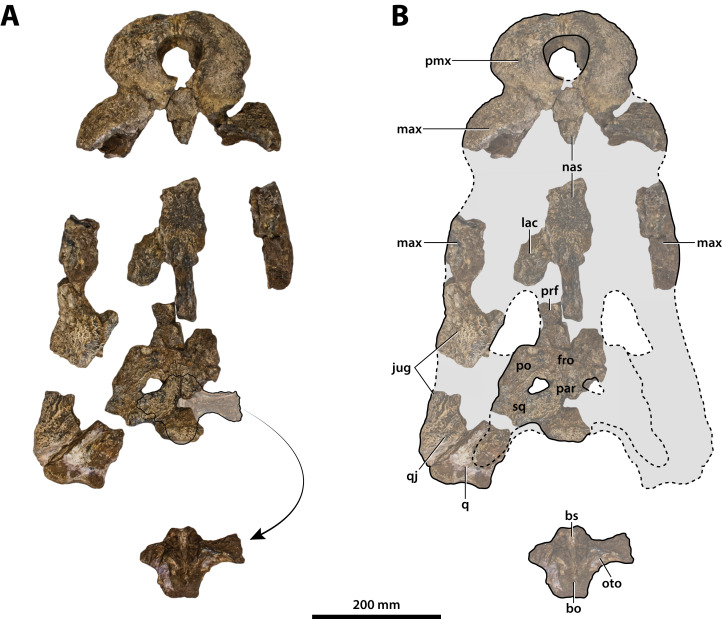
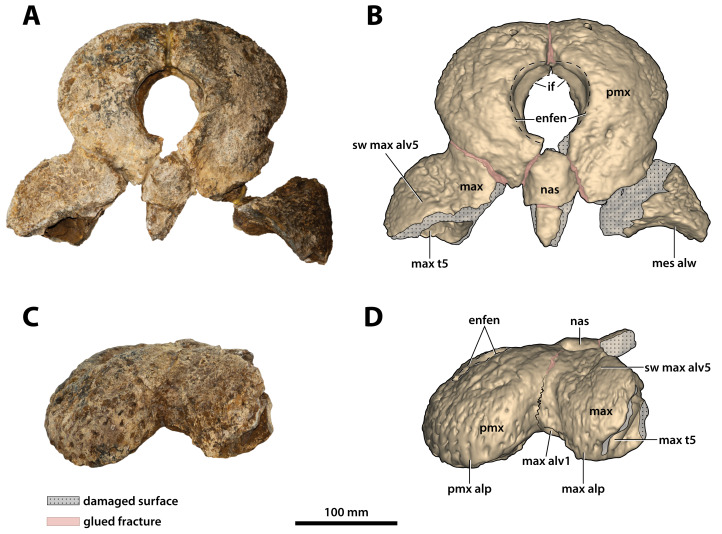
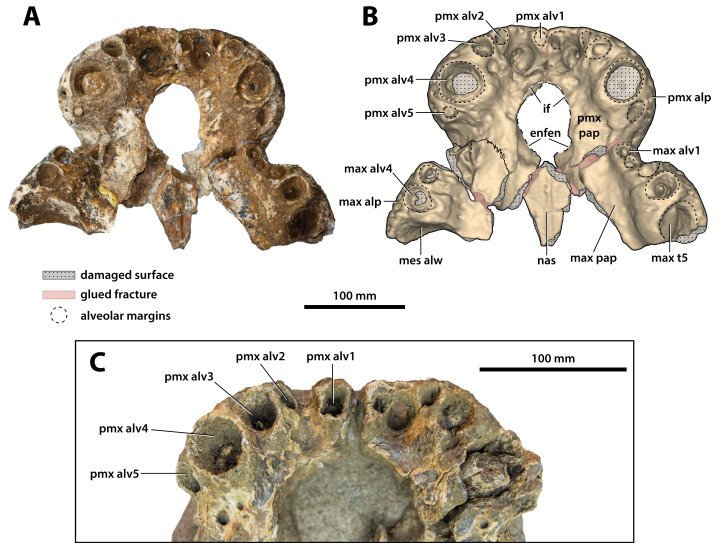
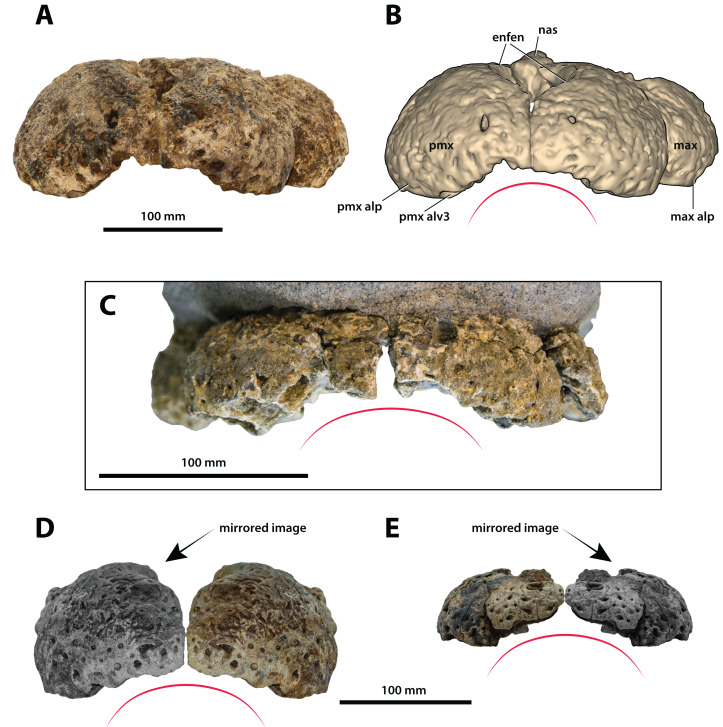
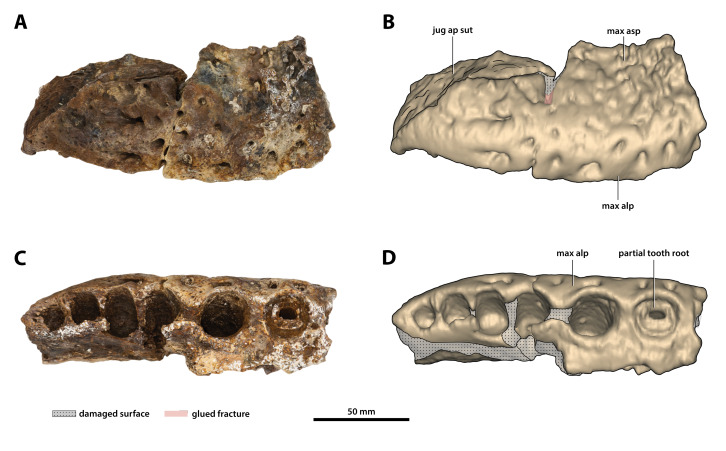
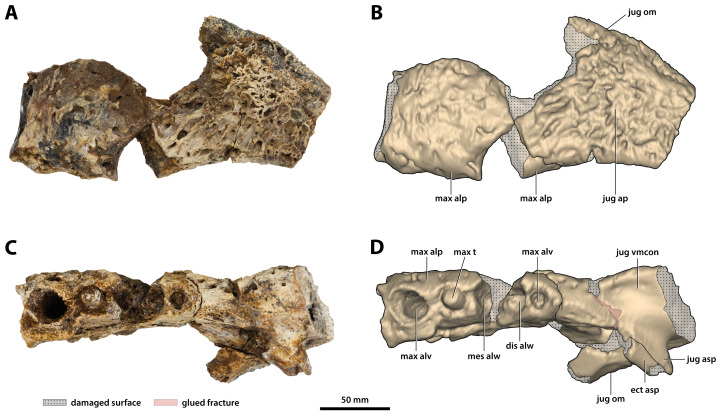
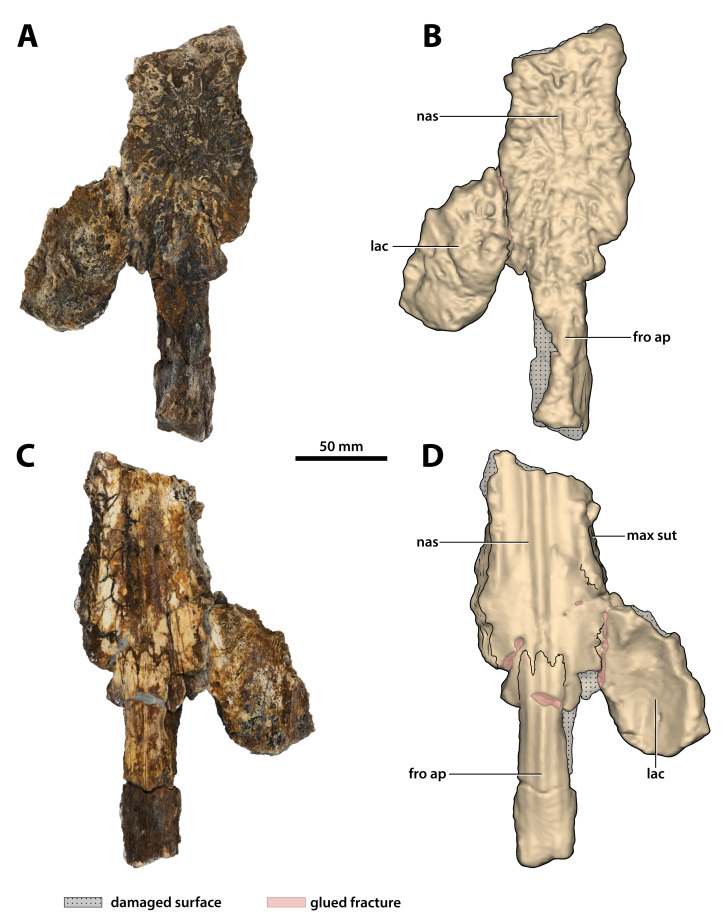
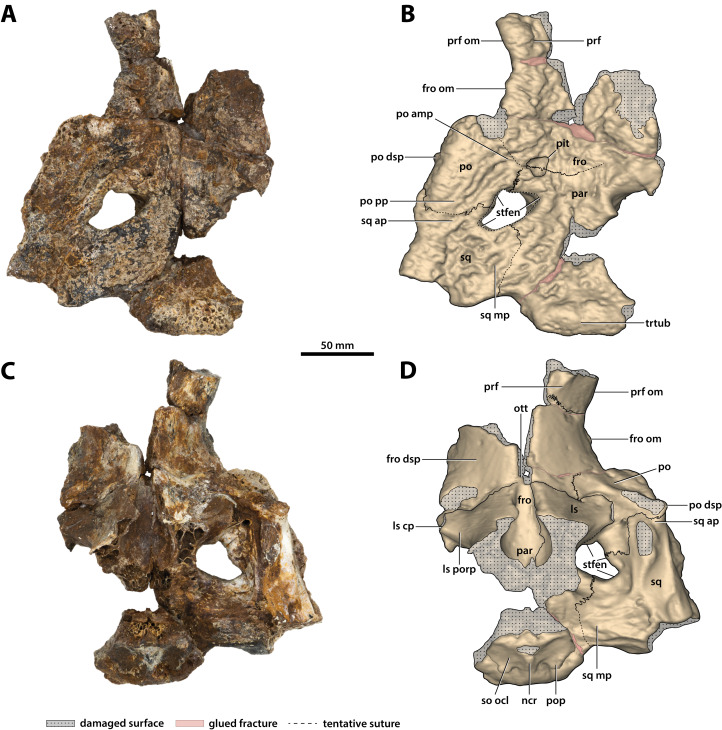
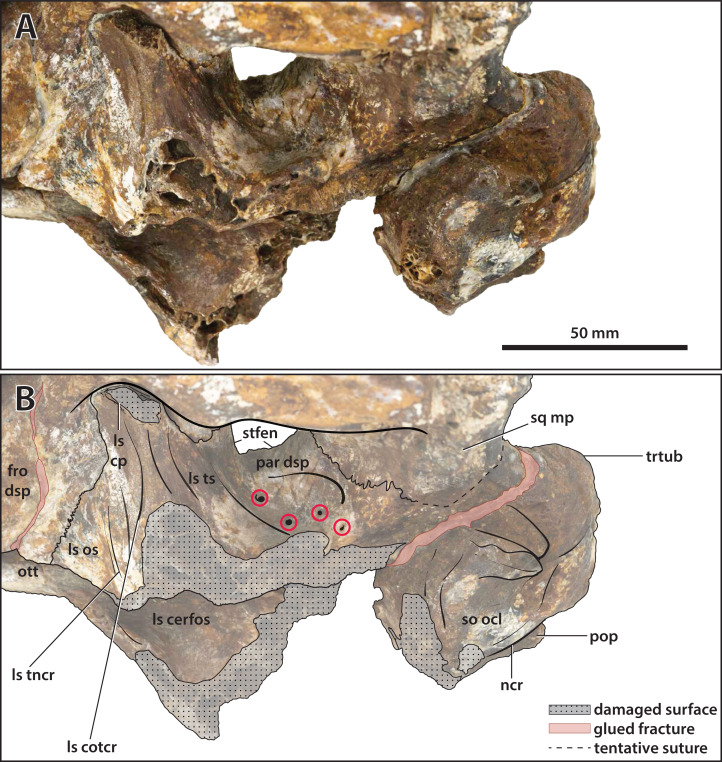
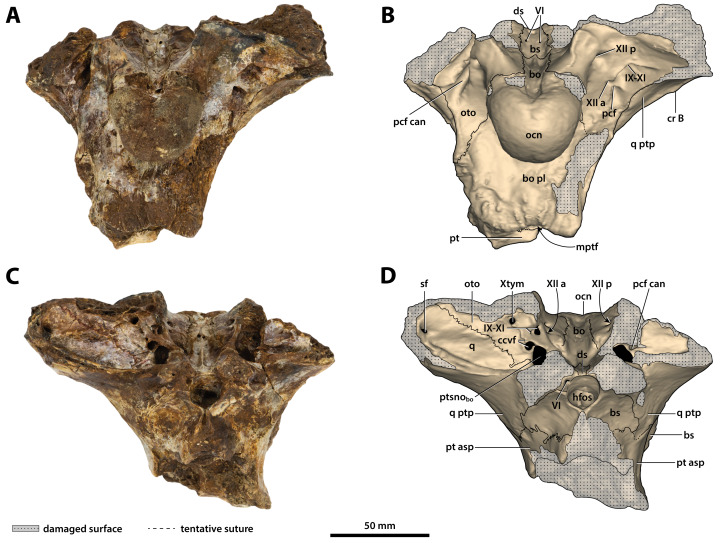
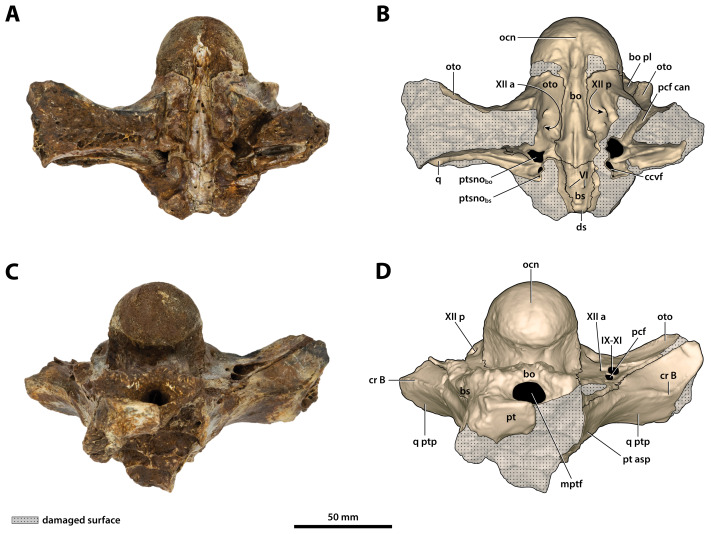
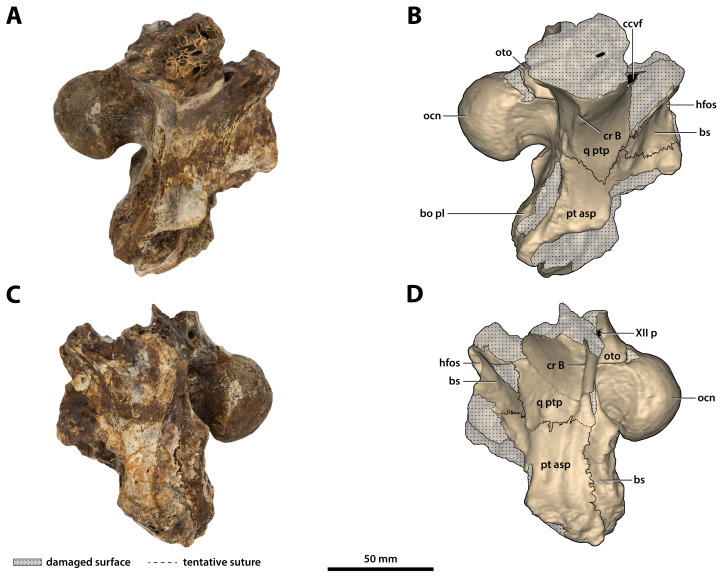
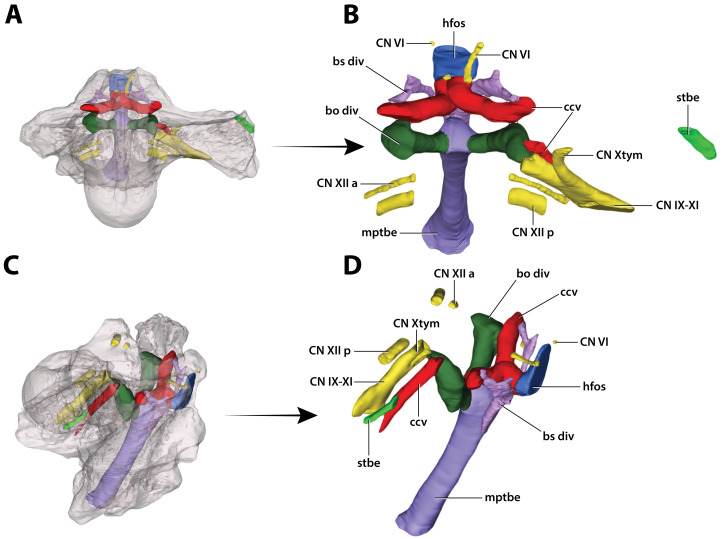
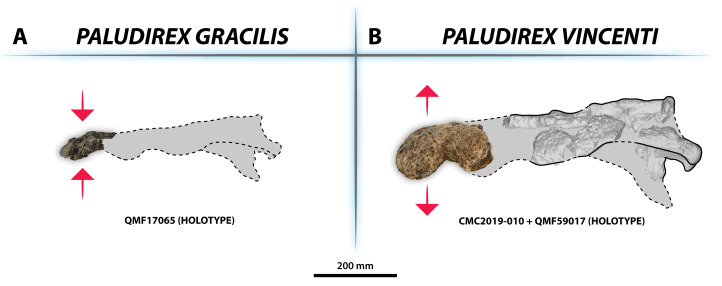
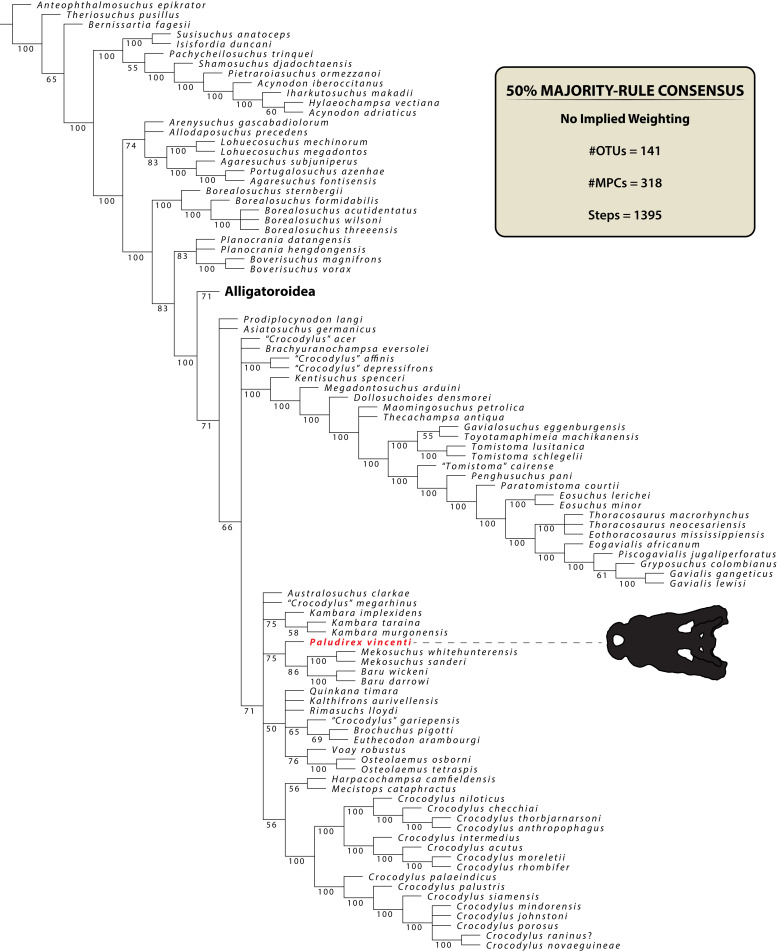
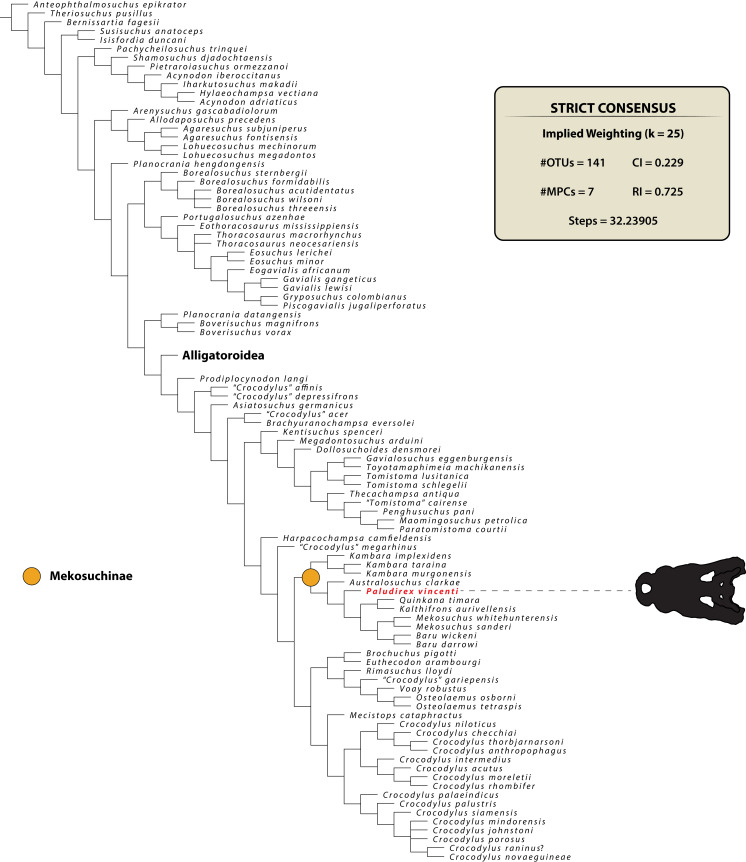
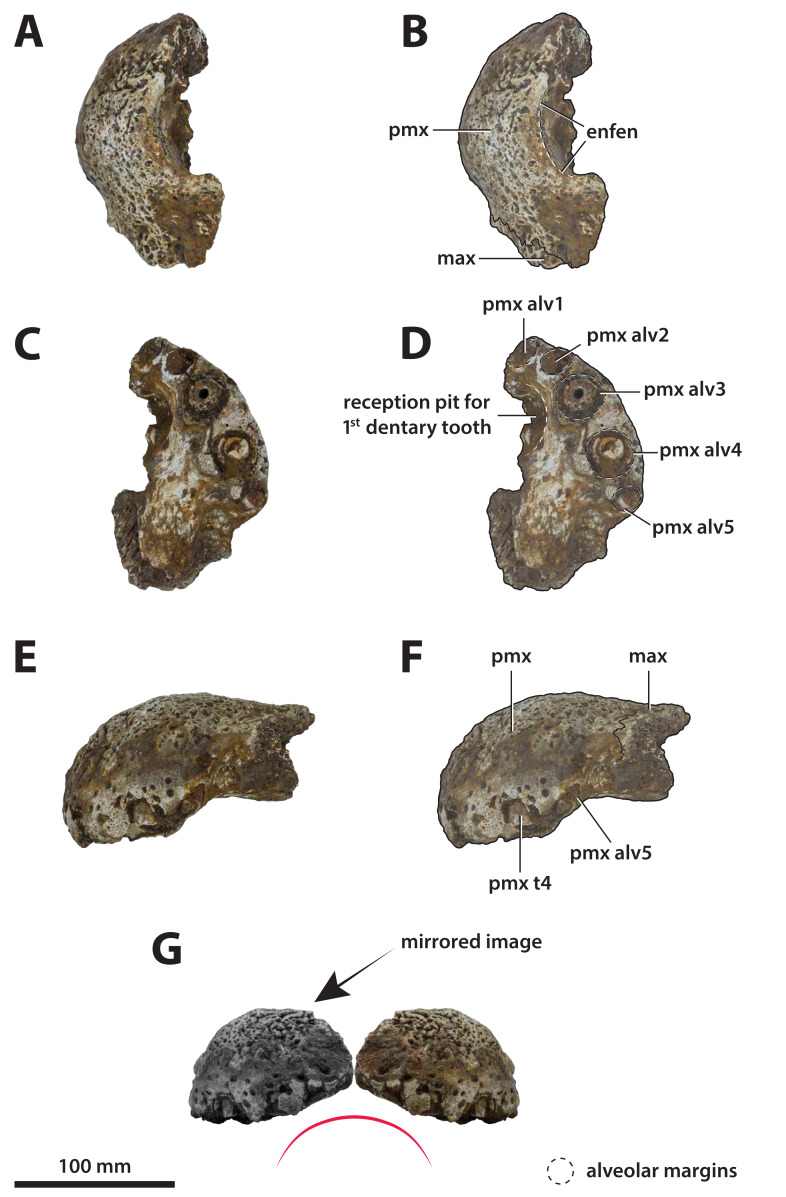
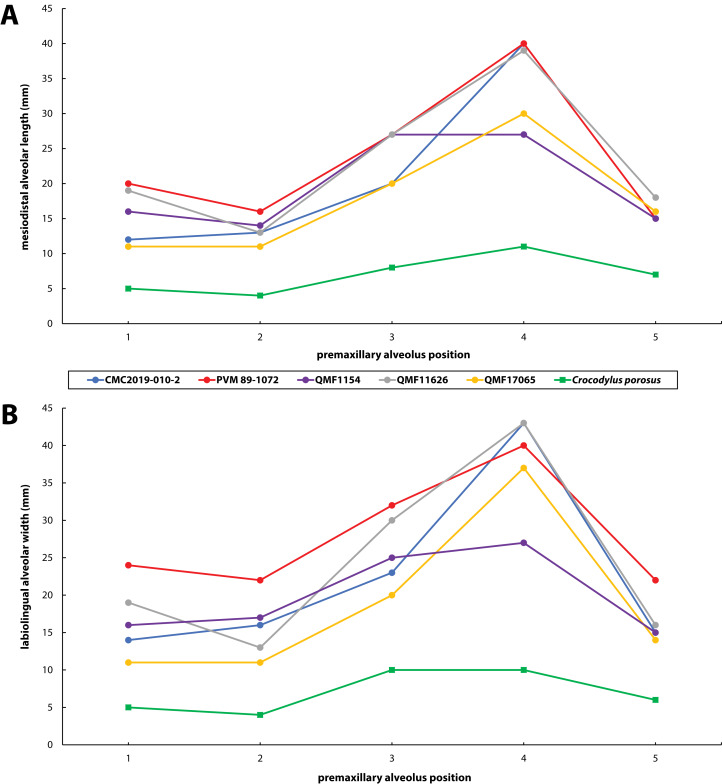
The crocodylian fossil record from the Cenozoic of Australasia is notable for its rich taxonomic diversity, and is primarily represented by members of the clade Mekosuchinae. Reports of crocodylian fossils from Australia date back to the late nineteenth century. In 1886, Charles Walter de Vis proposed the name for crocodylian fossils from southeast Queensland-the first binomen given to an extinct crocodylian taxon from Australia. has come to be regarded as a large, broad-snouted crocodylian from Australia's Plio-Pleistocene, and numerous specimens, few of which are sufficiently complete, have been assigned to it by several authors throughout the twentieth century. In the late 1990s, the genus was expanded to include a second species, . Unfortunately, the original syntype series described as is very fragmentary and derives from more than one taxon, while a large part of the subsequently selected lectotype specimen is missing. Because descriptions and illustrations of the complete lectotype do not reveal any autapomorphic features, we propose that should be regarded as a . Following this decision, the fossil material previously referred to is of uncertain taxonomic placement. A partial skull, formerly assigned to and known as 'Geoff Vincent's specimen', possesses many features of diagnostic value and is therefore used as basis to erect a new genus and species gen. et sp. nov. A comprehensive description is given for the osteology of 'Geoff Vincent's specimen' as well as aspects of its palaeoneurology, the latter being a first for an extinct Australian crocodyliform. The newly named genus is characterized by a unique combination of premaxillary features such as a distinctive arching of the anterior alveolar processes of the premaxillae, a peculiar arrangement of the first two premaxillary alveoli and a large size disparity between the 3rd and 4th premaxillary alveoli. These features presently allow formal recognition of two species within the genus, and comb. nov., with the former having comparatively more robust rostral proportions than the latter. The holotype comes from the Pliocene Chinchilla Sand of the Darling Downs, south-eastern Queensland, whereas the material assigned to is from the Pleistocene of Terrace Site Local Fauna, Riversleigh, northwest Queensland. Phylogenetic analyses recover as a mekosuchine, although further cladistic assessments are needed to better understand the relationships within the clade.
澳大拉西亚新生代的鳄鱼化石记录以其丰富的分类多样性而闻名,主要由麦氏鳄亚科的成员代表。澳大利亚鳄鱼化石的报告可追溯到19世纪末。1886年,查尔斯·沃尔特·德·维斯为昆士兰东南部的鳄鱼化石命名,这是给澳大利亚已灭绝鳄鱼分类单元的第一个双名法名称。后来被认为是来自澳大利亚上新世 - 更新世的一种大型宽吻鳄鱼,在整个20世纪,许多作者将众多标本(其中很少有足够完整的)归入该属。在20世纪90年代后期,该属被扩大到包括第二个物种。不幸的是,最初描述为的模式系列非常破碎,且来自不止一个分类单元,而随后选定的主模式标本的很大一部分缺失了。由于完整主模式标本的描述和图示未揭示任何自近裔性状特征,我们提议应将视为一个。基于这一决定,先前归入的化石材料的分类位置不确定。一个部分头骨,以前归入并被称为“杰夫·文森特标本”,具有许多诊断价值的特征,因此被用作建立一个新属和新物种的基础。对“杰夫·文森特标本”的骨骼学以及古神经学方面进行了全面描述(后者是对已灭绝澳大利亚鳄形类动物的首次描述)。新命名的属的特征是上颌骨特征的独特组合,如下颌前齿槽突的独特拱形、前两个上颌齿槽的特殊排列以及第三和第四上颌齿槽之间较大的尺寸差异。目前,这些特征使得该属内能够正式识别出两个物种,和新组合,前者的吻部比例相对比后者更粗壮。的正模标本来自昆士兰东南部达令唐斯的上新世钦奇拉砂岩,而归入的材料来自昆士兰西北部里弗斯利阶地遗址地方动物群的更新世。系统发育分析将恢复为麦氏鳄类,尽管需要进一步的分支系统评估来更好地理解该类群内部的关系。