ULB Neuroscience Institute (UNI), Université Libre de Bruxelles (ULB), B-6041, Gosselies, Belgium.
Paris-Saclay Institute of Neuroscience, CNRS, Univ Paris Sud, Université Paris-Saclay, UMR 9197- Neuro-PSI, Bat. 445, 91405, ORSAY Cedex, France.
Neural Dev. 2017 Sep 1;12(1):16. doi: 10.1186/s13064-017-0093-2.
Amacrine interneurons that modulate synaptic plasticity between bipolar and ganglion cells constitute the most diverse cell type in the retina. Most are inhibitory neurons using either GABA or glycine as neurotransmitters. Although several transcription factors involved in amacrine cell fate determination have been identified, mechanisms underlying amacrine cell subtype specification remain to be further understood. The Prdm13 histone methyltransferase encoding gene is a target of the transcription factor Ptf1a, an essential regulator of inhibitory neuron cell fate in the retina. Here, we have deepened our knowledge on its interaction with Ptf1a and investigated its role in amacrine cell subtype determination in the developing Xenopus retina.
We performed prdm13 gain and loss of function in Xenopus and assessed the impact on retinal cell fate determination using RT-qPCR, in situ hybridization and immunohistochemistry.
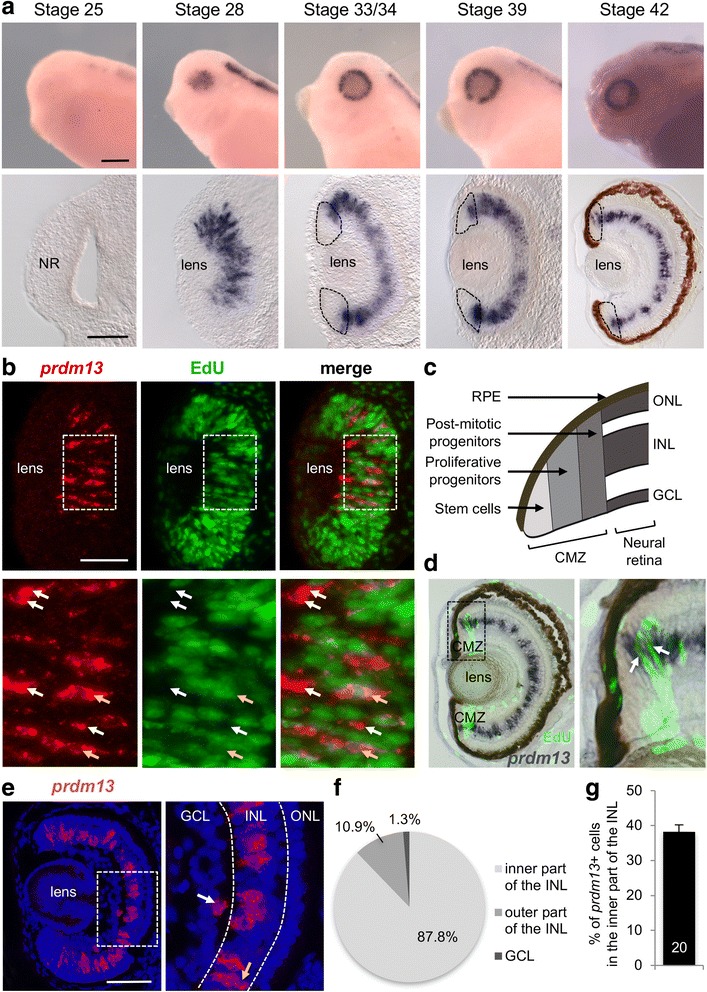
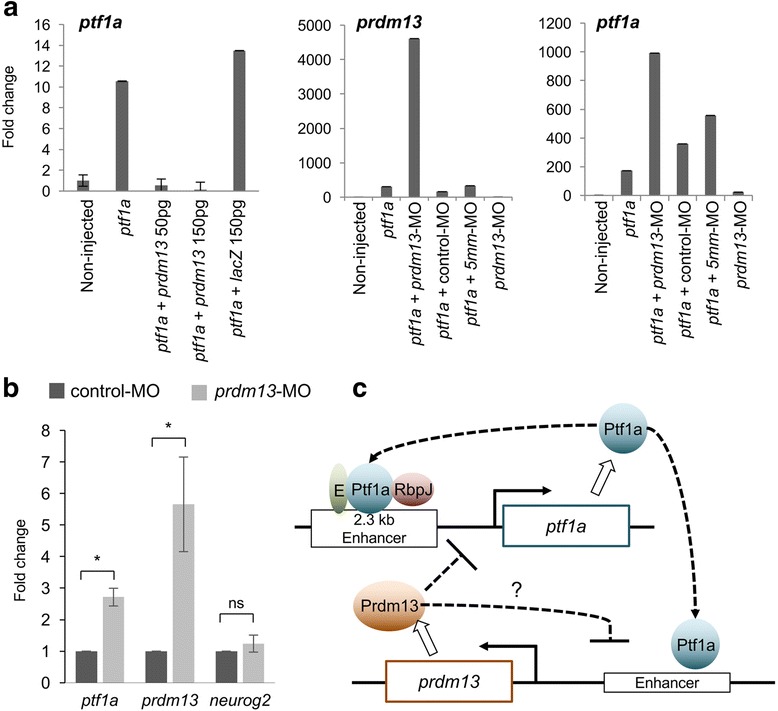
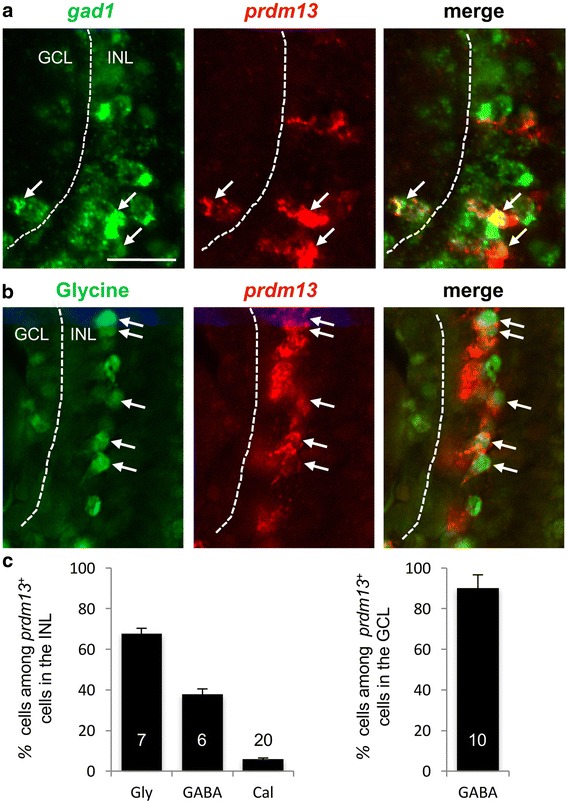
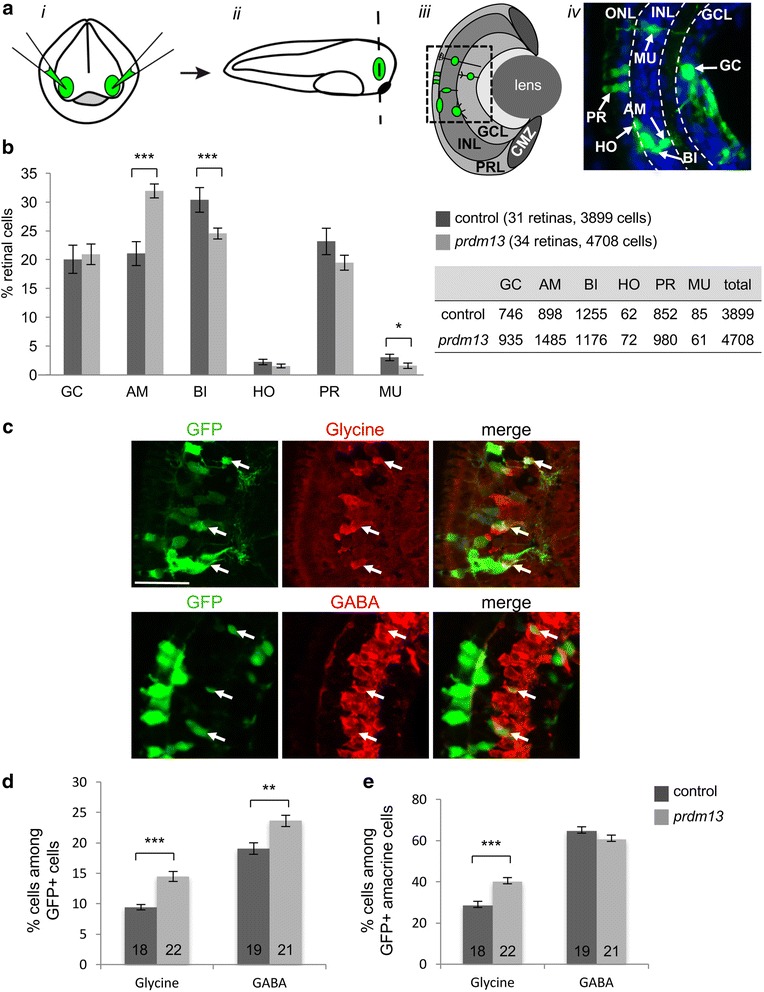
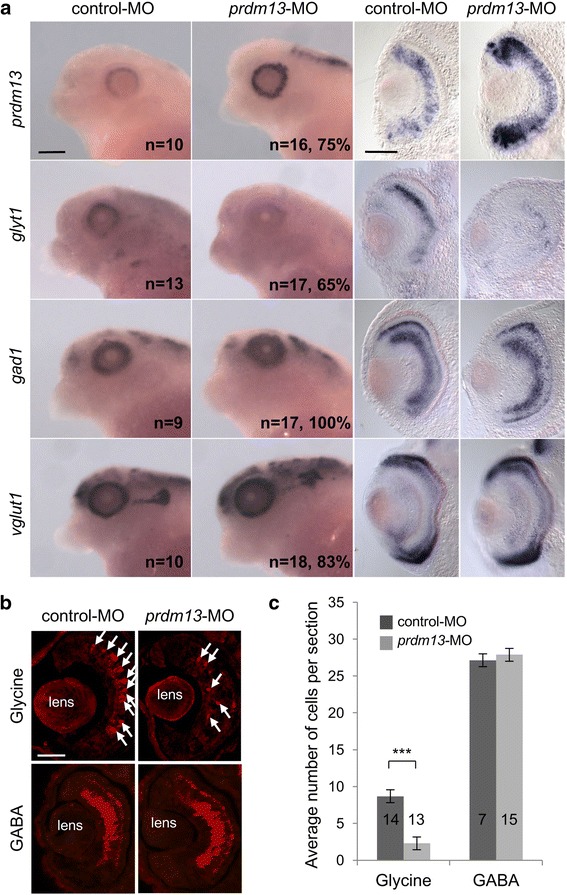
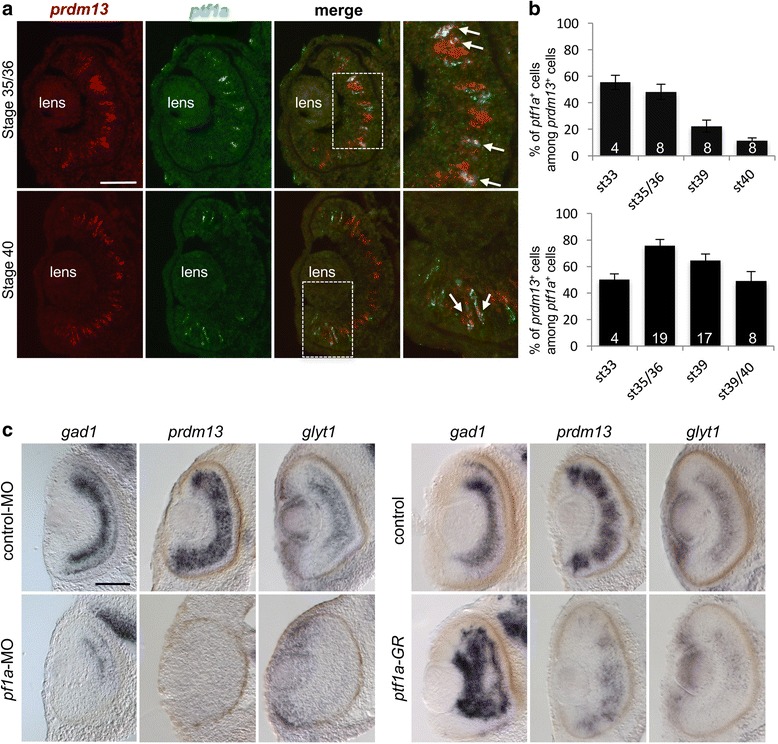
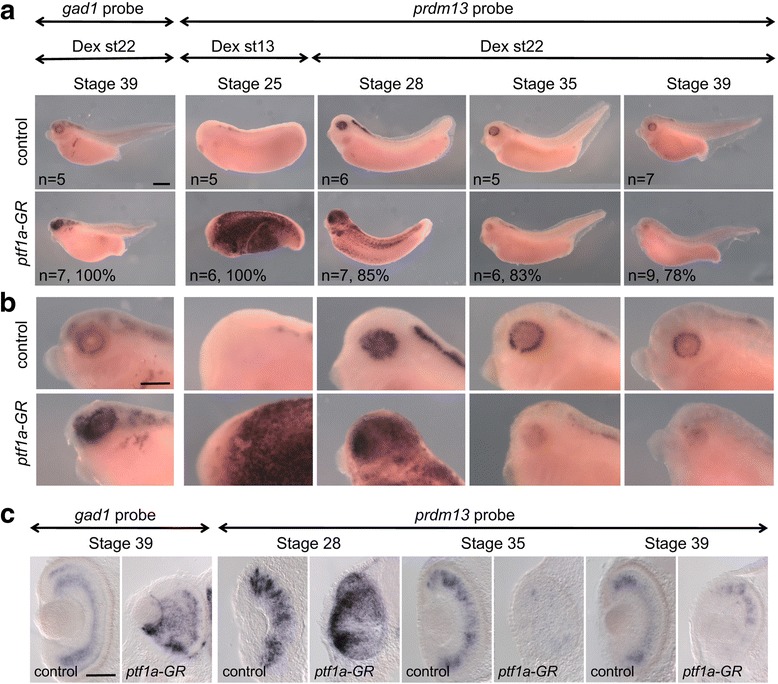
We found that prdm13 in the amphibian Xenopus is expressed in few retinal progenitors and in about 40% of mature amacrine cells, predominantly in glycinergic ones. Clonal analysis in the retina reveals that prdm13 overexpression favours amacrine cell fate determination, with a bias towards glycinergic cells. Conversely, knockdown of prdm13 specifically inhibits glycinergic amacrine cell genesis. We also showed that, as in the neural tube, prdm13 is subjected to a negative autoregulation in the retina. Our data suggest that this is likely due to its ability to repress the expression of its inducer, ptf1a.
Our results demonstrate that Prdm13, downstream of Ptf1a, acts as an important regulator of glycinergic amacrine subtype specification in the Xenopus retina. We also reveal that Prdm13 regulates ptf1a expression through a negative feedback loop.
调节双极细胞和节细胞之间突触可塑性的无长突细胞是视网膜中最多样化的细胞类型。大多数为抑制性神经元,使用 GABA 或甘氨酸作为神经递质。尽管已经确定了几个参与无长突细胞命运决定的转录因子,但无长突细胞亚型特化的机制仍有待进一步了解。Prdm13 组蛋白甲基转移酶编码基因是转录因子 Ptf1a 的靶标,Ptf1a 是视网膜抑制性神经元命运的必需调节因子。在这里,我们深入了解了它与 Ptf1a 的相互作用,并研究了它在发育中的非洲爪蟾视网膜中无长突细胞亚型决定中的作用。
我们在非洲爪蟾中进行了 prdm13 的功能获得和功能丧失,并使用 RT-qPCR、原位杂交和免疫组织化学评估了其对视网膜细胞命运决定的影响。
我们发现,两栖动物非洲爪蟾中的 prdm13 在少数视网膜祖细胞和大约 40%的成熟无长突细胞中表达,主要在甘氨酸能无长突细胞中。视网膜克隆分析显示,prdm13 过表达有利于无长突细胞命运的决定,偏向于甘氨酸能细胞。相反,prdm13 的敲低特异性抑制甘氨酸能无长突细胞的发生。我们还表明,与神经管一样,prdm13 在视网膜中受到负自调控。我们的数据表明,这可能是由于其能够抑制其诱导物 ptf1a 的表达。
我们的结果表明,Prdm13 是 Ptf1a 的下游因子,作为非洲爪蟾视网膜中甘氨酸能无长突细胞亚型特化的重要调节因子。我们还揭示了 Prdm13 通过负反馈环调节 ptf1a 的表达。