Cedano Prieto Dora Maria, Cheng Yushen, Chang Chih-Chieh, Yu Jessica, Takada Yoko K, Takada Yoshikazu
Department of Dermatology, Biochemistry and Molecular Medicine, UC Davis School of Medicine, Research III Suite, 4645 Second Avenue, Sacramento, CA, United States of America.
College of Medical Science and Technology, Taipei Medical University, Taipei, Taiwan.
PLoS One. 2017 Sep 5;12(9):e0184285. doi: 10.1371/journal.pone.0184285. eCollection 2017.
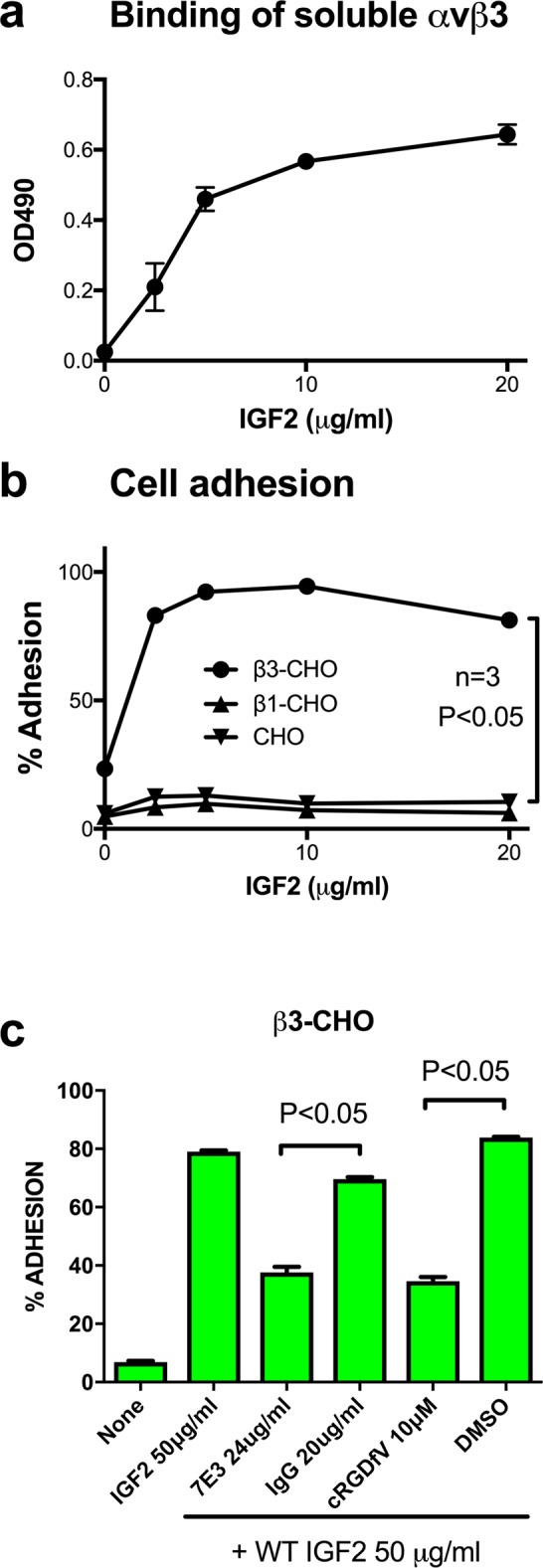
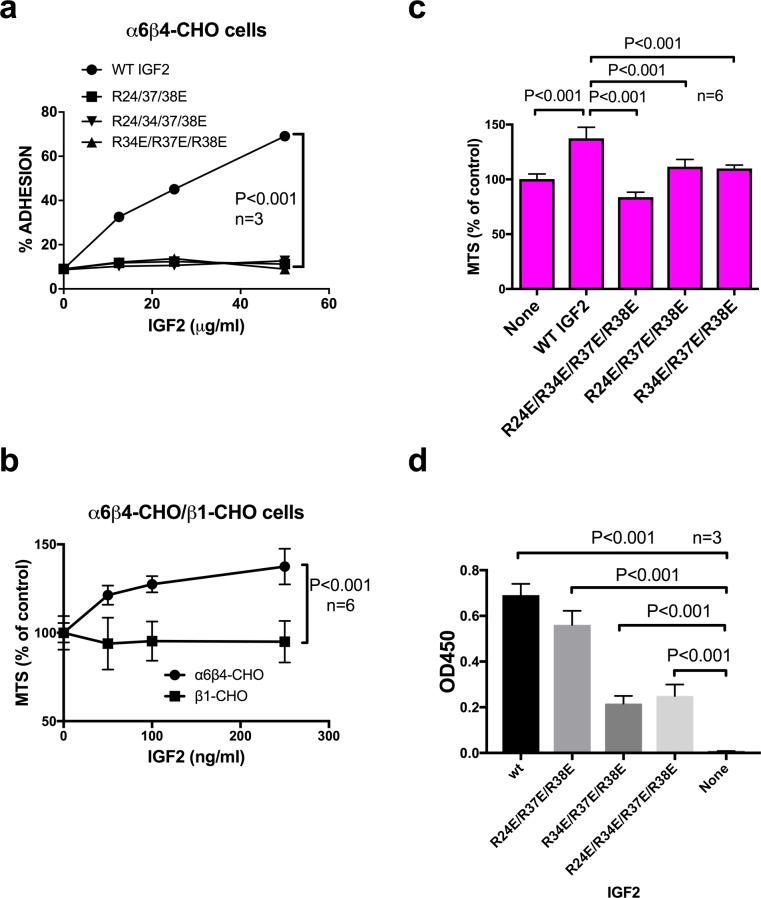
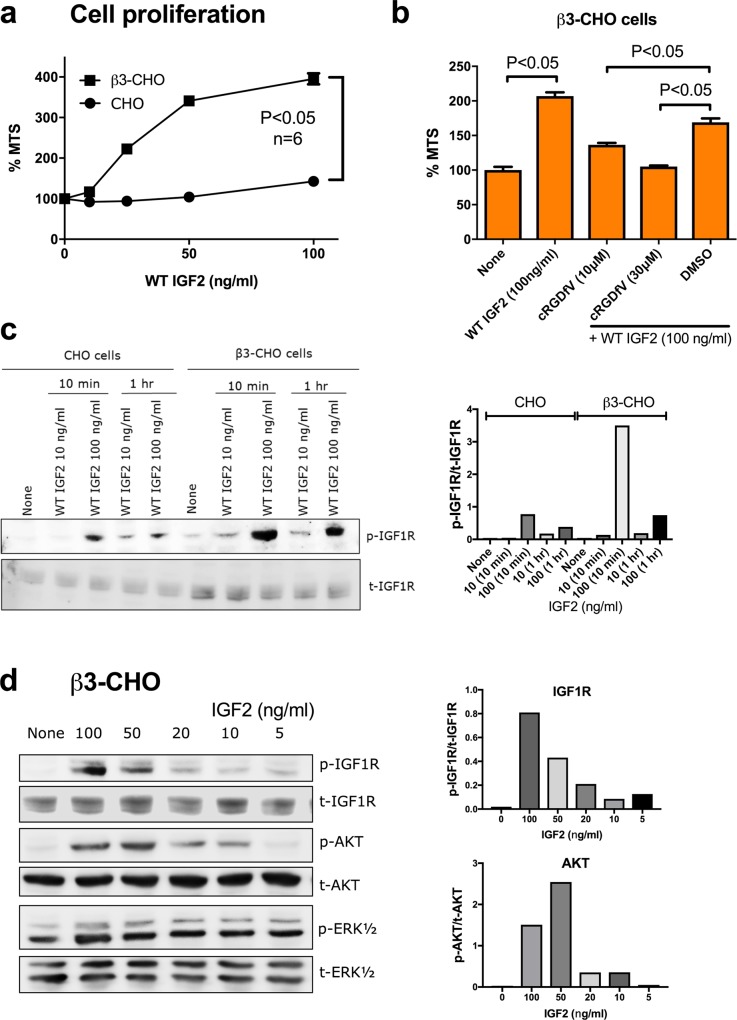
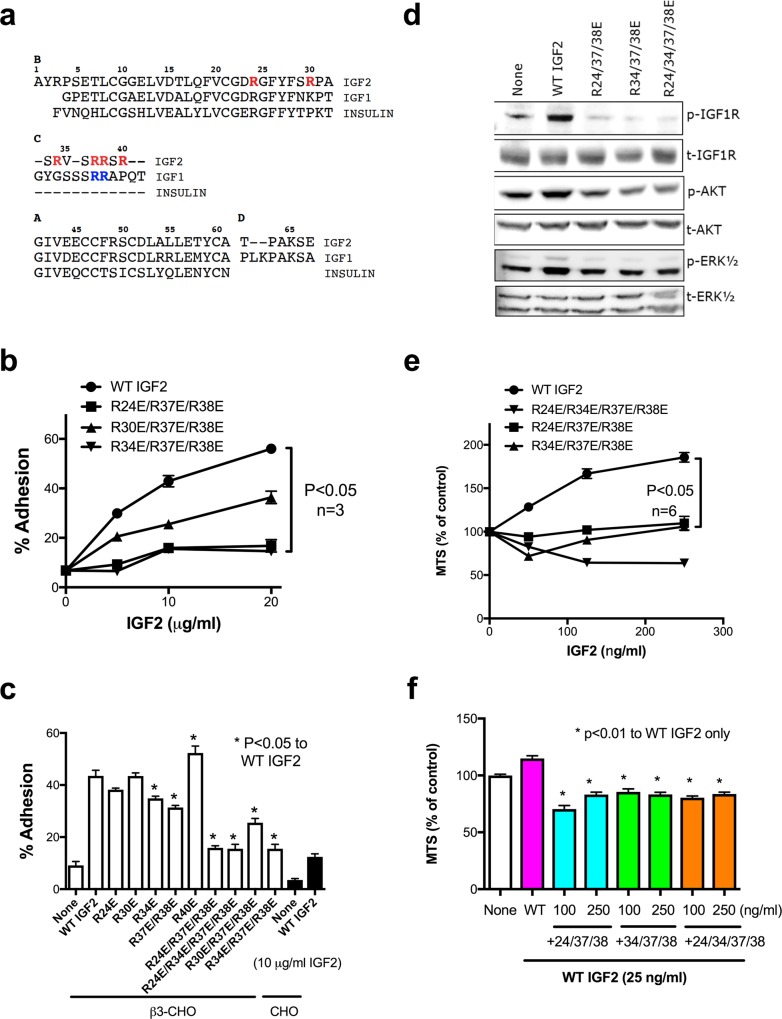
We have reported that integrins crosstalk with growth factors through direct binding to growth factors (e.g., fibroblast growth factor-1, insulin-like growth factor 1 (IGF1), neuregulin-1, fractalkine) and subsequent ternary complex formation with cognate receptor [e.g., integrin/IGF1/IGF1 receptor (IGF1R)]. IGF1 and IGF2 are overexpressed in cancer and major therapeutic targets. We previously reported that IGF1 binds to integrins ανβ3 and α6β4, and the R36E/R37E mutant in the C-domain of IGF1 is defective integrin binding and signaling functions of IGF1, and acts as an antagonist of IGF1R. We studied if integrins play a role in the signaling functions of IGF2, another member of the IGF family. Here we describe that IGF2 specifically binds to integrins ανβ3 and α6β4, and induced proliferation of CHO cells (IGF1R+) that express ανβ3 or α6β4 (β3- or α6β4-CHO cells). Arg residues to Glu at positions 24, 34, 37 and/or 38 in or close to the C-domain of IGF2 play a critical role in binding to integrins and signaling functions. The R24E/R37E/R38E, R34E/R37E/R38E, and R24E/R34E/R37E/R38E mutants were defective in integrin binding and IGF2 signaling. These mutants suppressed proliferation induced by WT IGF2, suggesting that they are dominant-negative antagonists of IGF1R. These results suggest that IGF2 also requires integrin binding for signaling functions, and the IGF2 mutants that cannot bind to integrins act as antagonists of IGF1R. The present study defines the role of the C-domain in integrin binding and signaling.
我们曾报道整合素通过直接结合生长因子(如成纤维细胞生长因子 -1、胰岛素样生长因子 1(IGF1)、神经调节蛋白 -1、趋化因子)与生长因子发生相互作用,并随后与同源受体形成三元复合物[如整合素/IGF1/IGF1 受体(IGF1R)]。IGF1 和 IGF2 在癌症中过表达,是主要的治疗靶点。我们之前报道过 IGF1 与整合素 ανβ3 和 α6β4 结合,IGF1 C 结构域中的 R36E/R37E 突变体在整合素结合及 IGF1 的信号传导功能方面存在缺陷,且可作为 IGF1R 的拮抗剂。我们研究了整合素是否在 IGF 家族的另一个成员 IGF2 的信号传导功能中发挥作用。在此我们描述,IGF2 特异性结合整合素 ανβ3 和 α6β4,并诱导表达 ανβ3 或 α6β4 的 CHO 细胞(IGF1R +)(β3 - 或 α6β4 - CHO 细胞)增殖。IGF2 C 结构域内或附近第 24、34、37 和/或 38 位的精氨酸残基突变为谷氨酸在整合素结合及信号传导功能中起关键作用。R24E/R37E/R38E、R34E/R37E/R38E 和 R24E/R34E/R37E/R38E 突变体在整合素结合及 IGF2 信号传导方面存在缺陷。这些突变体抑制了野生型 IGF2 诱导的增殖,表明它们是 IGF1R 的显性负性拮抗剂。这些结果表明,IGF2 的信号传导功能也需要整合素结合,且不能与整合素结合的 IGF2 突变体可作为 IGF1R 的拮抗剂。本研究确定了 C 结构域在整合素结合及信号传导中的作用。