Peter Medawar Building for Pathogen Research, Nuffield Department of Medicine, University of Oxford, Oxford, United Kingdom.
Biomedical Sciences Research Complex, University of St Andrews, St Andrews, United Kingdom.
Elife. 2017 Sep 29;6:e29112. doi: 10.7554/eLife.29112.
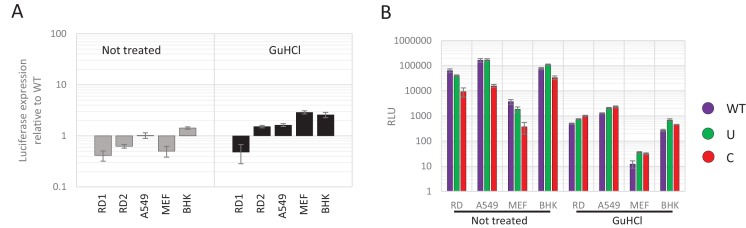
Most vertebrate and plant RNA and small DNA viruses suppress genomic CpG and UpA dinucleotide frequencies, apparently mimicking host mRNA composition. Artificially increasing CpG/UpA dinucleotides attenuates viruses through an entirely unknown mechanism. Using the echovirus 7 (E7) model in several cell types, we show that the restriction in E7 replication in mutants with increased CpG/UpA dinucleotides occurred immediately after viral entry, with incoming virions failing to form replication complexes. Sequences of CpG/UpA-high virus stocks showed no evidence of increased mutational errors that would render them replication defective, these viral RNAs were not differentially sequestered in cytoplasmic stress granules nor did they induce a systemic antiviral state. Importantly, restriction was not mediated through effects on translation efficiency since replicons with high CpG/UpA sequences inserted into a non-coding region were similarly replication defective. Host-cells thus possess intrinsic defence pathways that prevent replication of viruses with increased CpG/UpA frequencies independently of codon usage.
大多数脊椎动物和植物 RNA 病毒和小型 DNA 病毒会抑制基因组 CpG 和 UpA 二核苷酸频率,显然是在模仿宿主 mRNA 组成。人为增加 CpG/UpA 二核苷酸会通过一种完全未知的机制使病毒衰减。我们使用几种细胞类型中的肠病毒 7(E7)模型表明,在 CpG/UpA 二核苷酸增加的突变体中,E7 的复制限制发生在病毒进入后立即发生,进入的病毒颗粒未能形成复制复合物。CpG/UpA-高病毒株的序列没有证据表明增加的突变错误会使它们失去复制能力,这些病毒 RNA 没有在细胞质应激颗粒中被差异隔离,也没有诱导全身性抗病毒状态。重要的是,限制不是通过对翻译效率的影响来介导的,因为插入非编码区的高 CpG/UpA 序列的复制子也具有相同的复制缺陷。因此,宿主细胞具有内在的防御途径,可以防止 CpG/UpA 频率增加的病毒复制,而与密码子使用无关。