Department of Molecular and Cell Biology, University of California, Berkeley, CA, United States.
Department of Integrative Biology, University of California, Berkeley, CA, United States.
Elife. 2017 Nov 17;6:e29550. doi: 10.7554/eLife.29550.
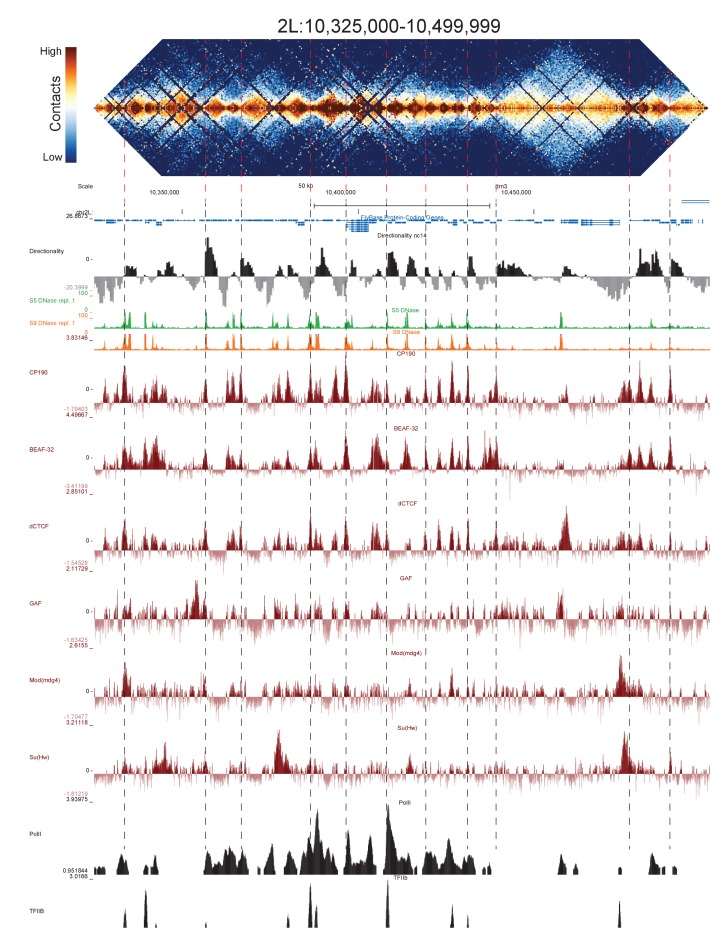
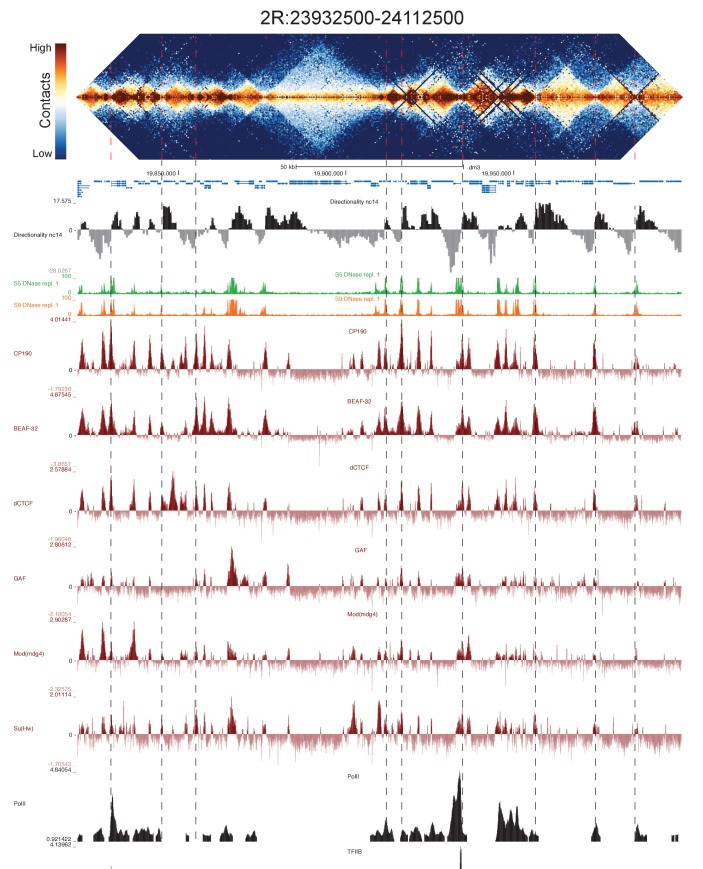
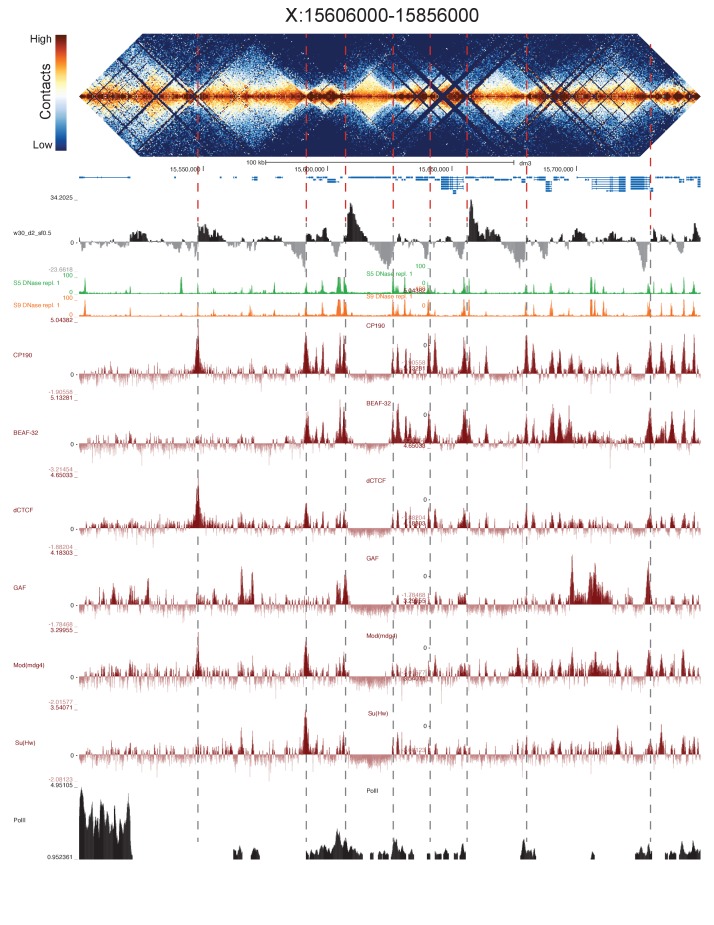
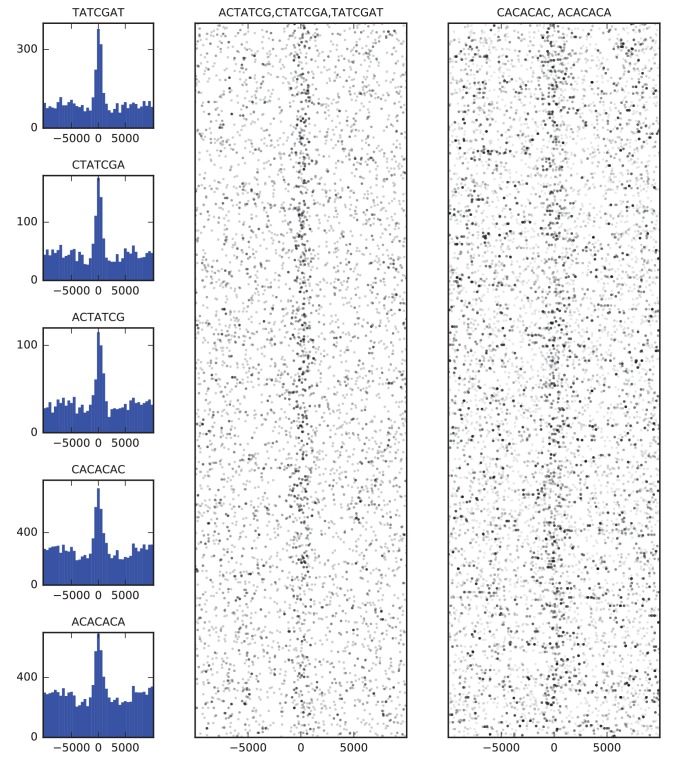
High-throughput assays of three-dimensional interactions of chromosomes have shed considerable light on the structure of animal chromatin. Despite this progress, the precise physical nature of observed structures and the forces that govern their establishment remain poorly understood. Here we present high resolution Hi-C data from early embryos. We demonstrate that boundaries between topological domains of various sizes map to DNA elements that resemble classical insulator elements: short genomic regions sensitive to DNase digestion that are strongly bound by known insulator proteins and are frequently located between divergent promoters. Further, we show a striking correspondence between these elements and the locations of mapped polytene interband regions. We believe it is likely this relationship between insulators, topological boundaries, and polytene interbands extends across the genome, and we therefore propose a model in which decompaction of boundary-insulator-interband regions drives the organization of interphase chromosomes by creating stable physical separation between adjacent domains.
高通量分析染色体的三维相互作用为动物染色质的结构提供了重要线索。尽管取得了这些进展,但观察到的结构的精确物理性质以及控制其形成的力仍未得到很好的理解。在这里,我们展示了来自早期胚胎的高分辨率 Hi-C 数据。我们证明,不同大小的拓扑域之间的边界映射到类似于经典绝缘子元件的 DNA 元件:对 DNA 酶消化敏感的短基因组区域,由已知的绝缘子蛋白强烈结合,并且经常位于发散启动子之间。此外,我们还展示了这些元件与映射的多线染色体间带区的位置之间存在惊人的对应关系。我们认为,绝缘子、拓扑边界和多线染色体间带之间的这种关系很可能在整个基因组中延伸,因此我们提出了一个模型,即边界-绝缘子-间带区域的解压缩通过在相邻域之间建立稳定的物理分离来驱动间期染色体的组织。