CNC - Center for Neuroscience and Cell Biology, University of Coimbra, Portugal; MIT-Portugal Program Bioengineering Systems Doctoral Program, Portugal.
CNC - Center for Neuroscience and Cell Biology, University of Coimbra, Portugal.
Redox Biol. 2018 May;15:297-315. doi: 10.1016/j.redox.2017.12.008. Epub 2017 Dec 21.
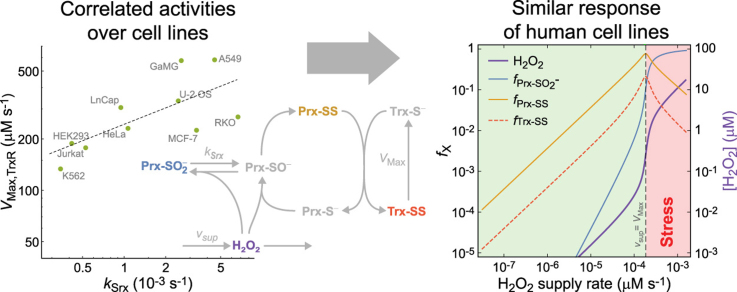
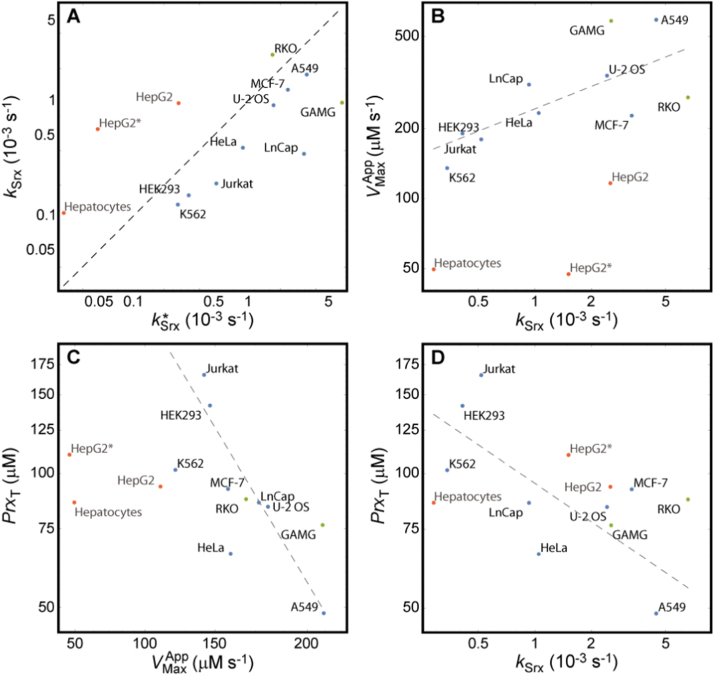
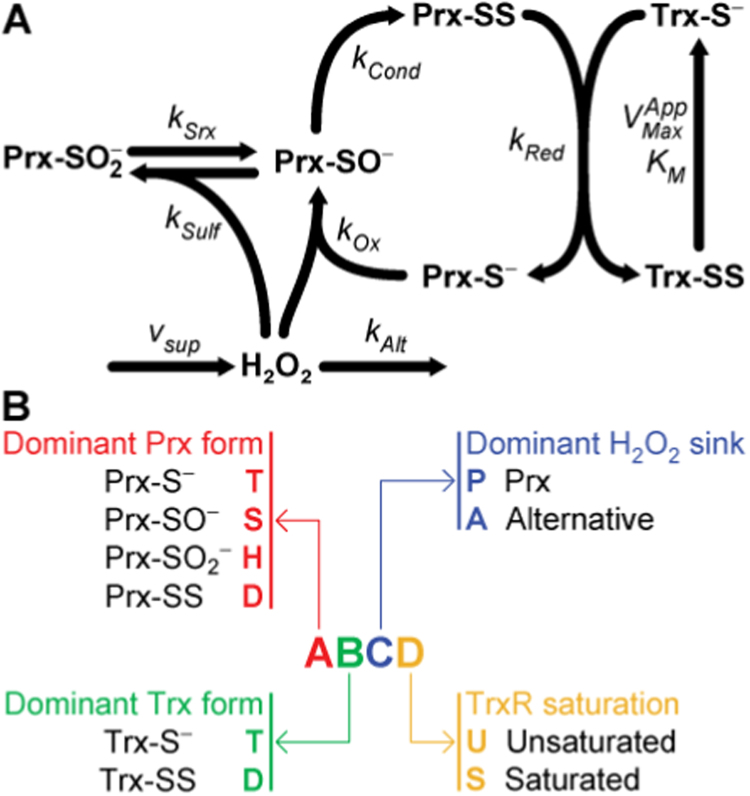
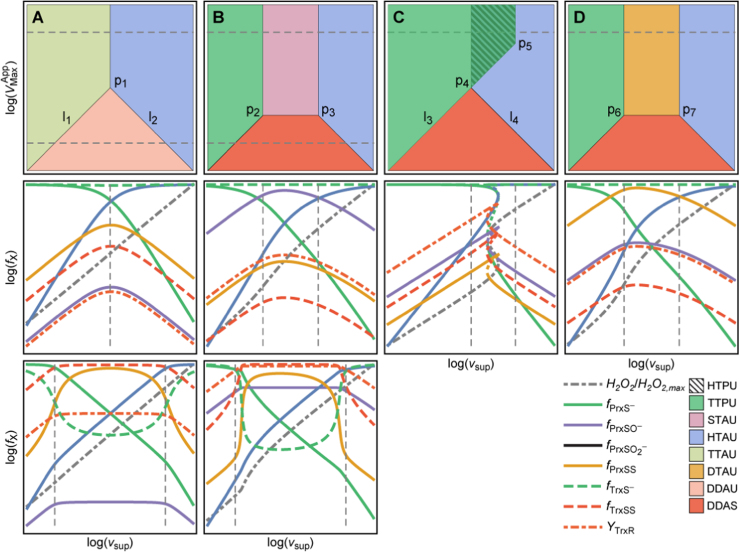
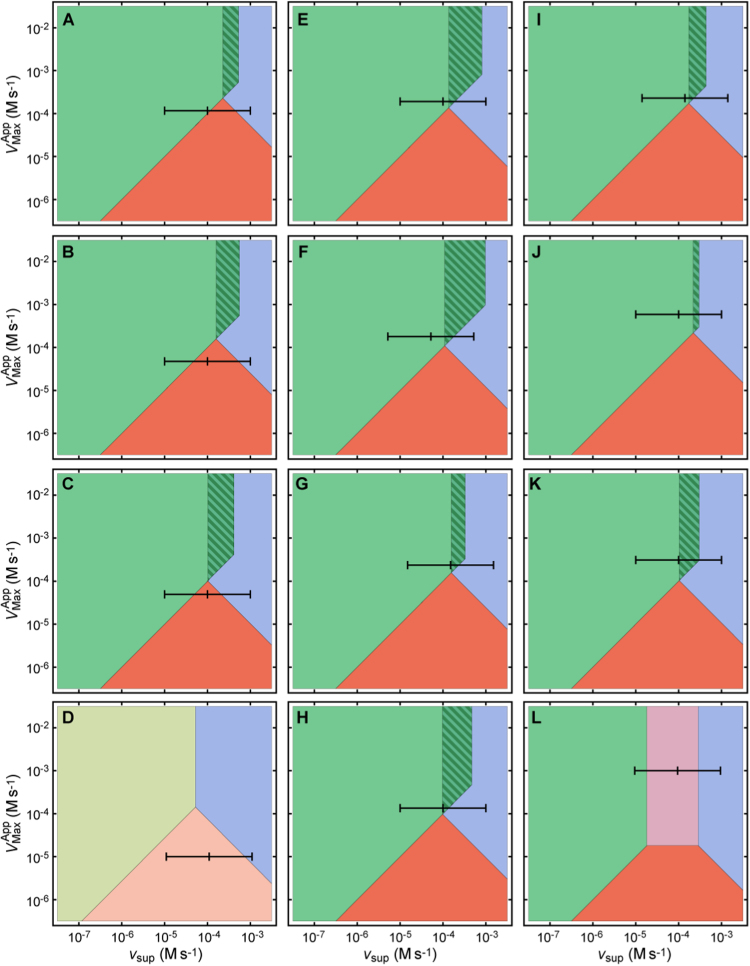
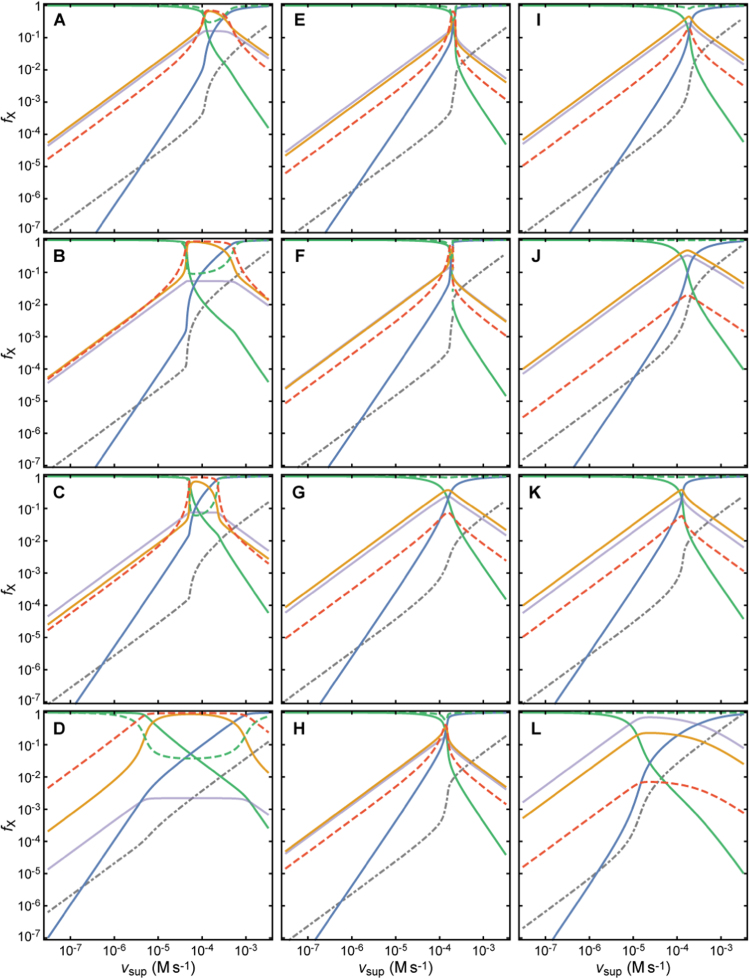
The system (PTTRS) formed by typical 2-Cys peroxiredoxins (Prx), thioredoxin (Trx), Trx reductase (TrxR), and sulfiredoxin (Srx) is central in antioxidant protection and redox signaling in the cytoplasm of eukaryotic cells. Understanding how the PTTRS integrates these functions requires tracing phenotypes to molecular properties, which is non-trivial. Here we analyze this problem based on a model that captures the PTTRS' conserved features. We have mapped the conditions that generate each distinct response to HO supply rates (v), and estimated the parameters for thirteen human cell types and for Saccharomyces cerevisiae. The resulting composition-to-phenotype map yielded the following experimentally testable predictions. The PTTRS permits many distinct responses including ultra-sensitivity and hysteresis. However, nearly all tumor cell lines showed a similar response characterized by limited Trx-S depletion and a substantial but self-limited gradual accumulation of hyperoxidized Prx at high v. This similarity ensues from strong correlations between the TrxR, Srx and Prx activities over cell lines, which contribute to maintain the Prx-SS reduction capacity in slight excess over the maximal steady state Prx-SS production. In turn, in erythrocytes, hepatocytes and HepG2 cells high v depletes Trx-S and oxidizes Prx mainly to Prx-SS. In all nucleated human cells the Prx-SS reduction capacity defined a threshold separating two different regimes. At sub-threshold v the cytoplasmic HO concentration is determined by Prx, nM-range and spatially localized, whereas at supra-threshold v it is determined by much less active alternative sinks and μM-range throughout the cytoplasm. The yeast shows a distinct response where the Prx Tsa1 accumulates in sulfenate form at high v. This is mainly due to an exceptional stability of Tsa1's sulfenate. The implications of these findings for thiol redox regulation and cell physiology are discussed. All estimates were thoroughly documented and provided, together with analytical approximations for system properties, as a resource for quantitative redox biology.
由典型的 2-Cys 过氧化物酶(Prx)、硫氧还蛋白(Trx)、Trx 还原酶(TrxR)和硫氧还蛋白(Srx)组成的系统(PTTRS)是真核细胞细胞质中抗氧化保护和氧化还原信号的核心。了解 PTTRS 如何整合这些功能需要追踪表型到分子特性,这并非易事。在这里,我们基于捕捉 PTTRS 保守特征的模型来分析这个问题。我们已经映射了生成每种不同的对 HO 供应速率(v)响应的条件,并为 13 个人类细胞类型和酿酒酵母估计了参数。由此产生的从组成到表型的映射产生了以下可通过实验验证的预测。PTTRS 允许许多不同的反应,包括超敏性和滞后性。然而,几乎所有的肿瘤细胞系都表现出相似的反应,其特征是 Trx-S 的耗竭有限,以及在高 v 时大量但自我限制的逐渐积累的过氧化 Prx。这种相似性是由于 TrxR、Srx 和 Prx 活性之间在细胞系之间存在强烈的相关性,这有助于保持 Prx-SS 还原能力略高于最大稳态 Prx-SS 产生。反过来,在红细胞、肝细胞和 HepG2 细胞中,高 v 会耗尽 Trx-S,并将 Prx 氧化主要为 Prx-SS。在所有有核人类细胞中,Prx-SS 还原能力定义了一个阈值,将两个不同的区域分开。在亚阈值 v 下,细胞质中的 HO 浓度由 Prx 决定,范围为 nM,空间上局限于 Prx;而在超阈值 v 下,它由细胞质中活性低得多的替代汇和范围为 μM 的物质决定。酵母表现出一种独特的反应,其中高 v 时 Prx Tsa1 以亚磺酸盐形式积累。这主要是由于 Tsa1 的亚磺酸盐稳定性异常高。这些发现对硫醇氧化还原调节和细胞生理学的意义将进行讨论。所有的估计都进行了彻底的记录,并与系统特性的分析近似值一起提供,作为定量氧化还原生物学的资源。