Department of Biology, The University of Iowa, Iowa City, Iowa 52242.
Department of Biology, The University of Iowa, Iowa City, Iowa 52242
Genetics. 2018 Apr;208(4):1421-1441. doi: 10.1534/genetics.118.300787. Epub 2018 Feb 22.
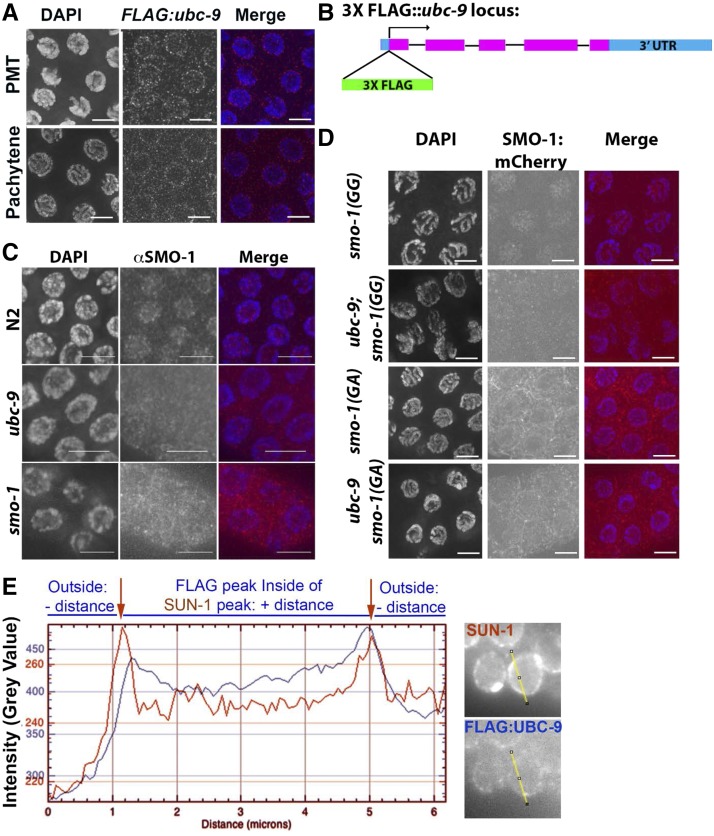
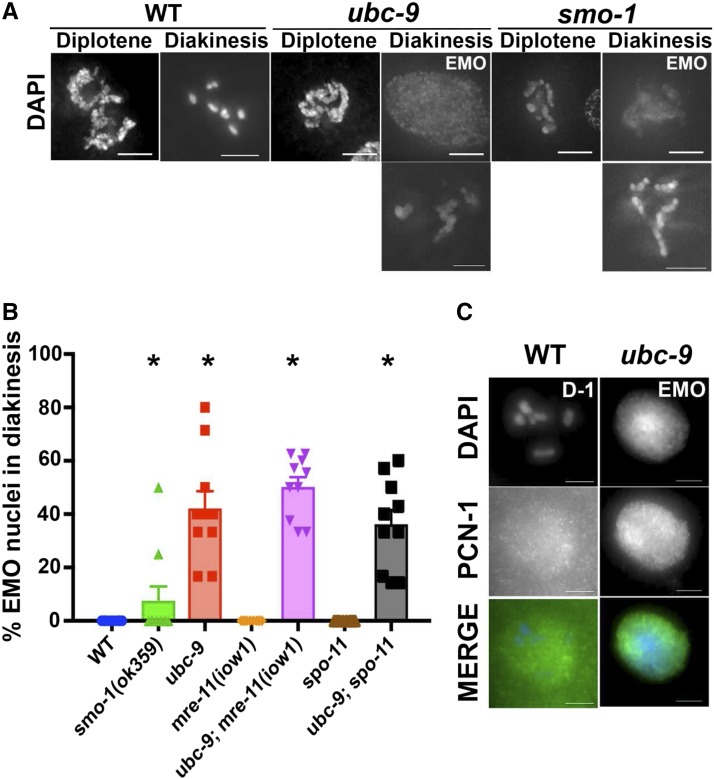
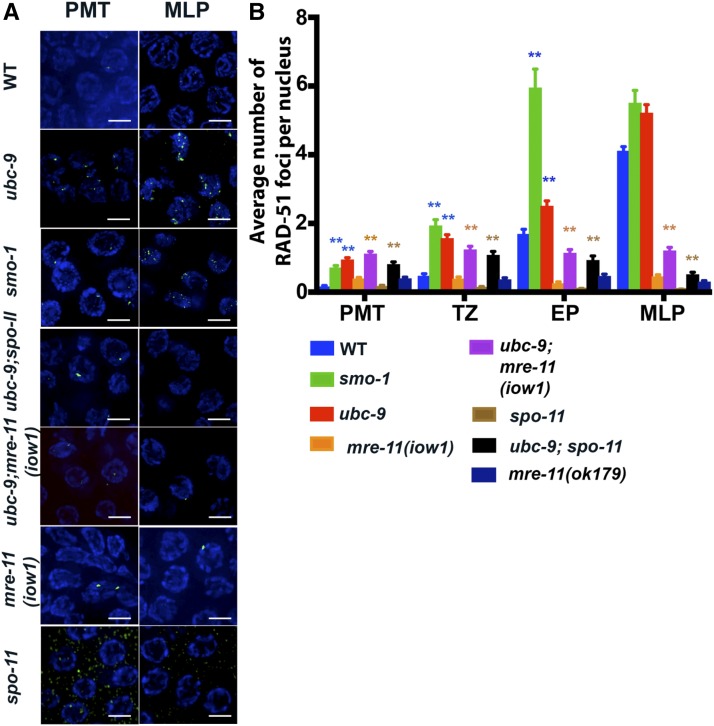
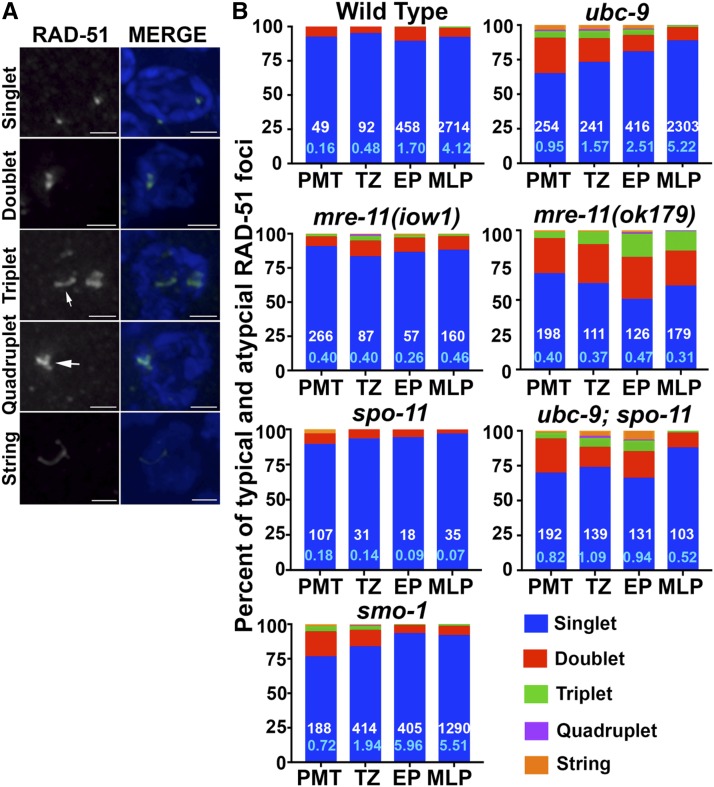
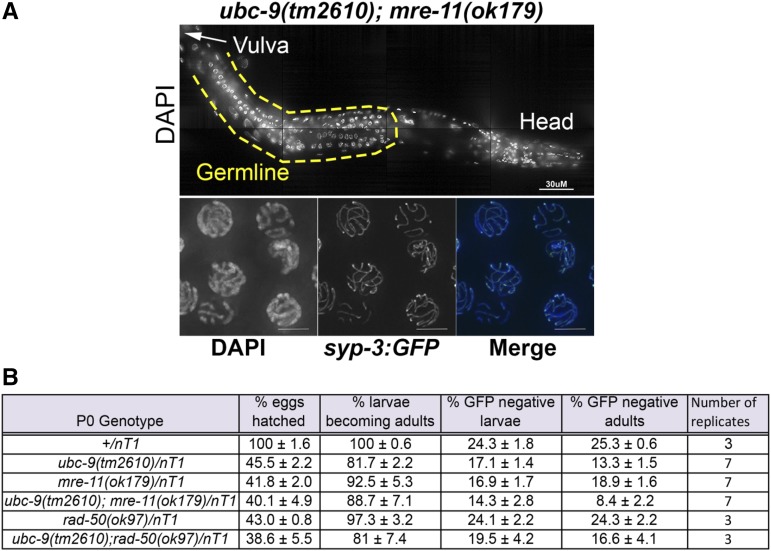
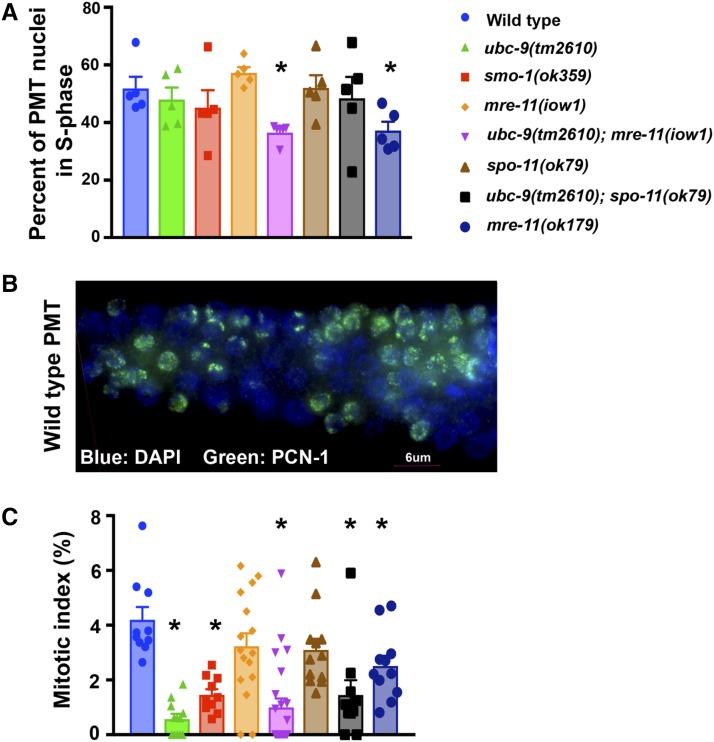
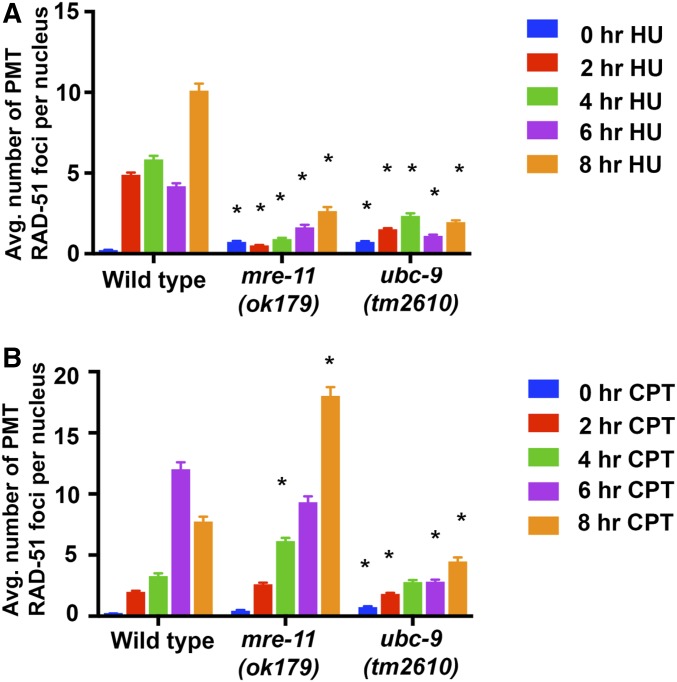
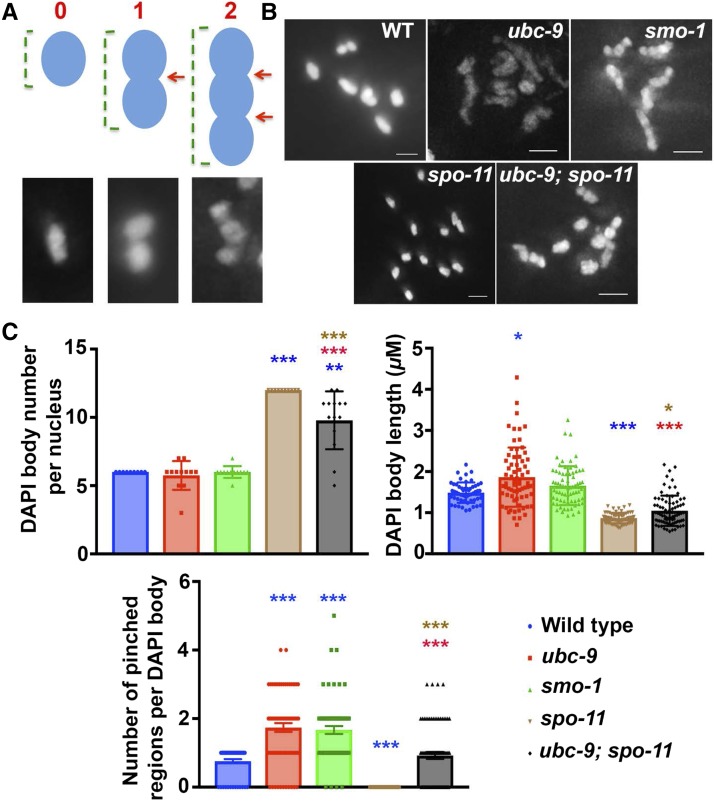
Meiosis is a highly regulated process, partly due to the need to break and then repair DNA as part of the meiotic program. Post-translational modifications are widely used during meiotic events to regulate steps such as protein complex formation, checkpoint activation, and protein attenuation. In this paper, we investigate how proteins that are obligatory components of the SUMO (small ubiquitin-like modifier) pathway, one such post-translational modification, affect the germline. We show that UBC-9, the E2 conjugation enzyme, and the homolog of SUMO, SMO-1, localize to germline nuclei throughout prophase I. Mutant analysis of and revealed increased recombination intermediates throughout the germline, originating during the mitotic divisions. SUMOylation mutants also showed late meiotic defects including defects in the restructuring of oocyte bivalents and endomitotic oocytes. Increased rates of noninterfering crossovers were observed in heterozygotes, even though interfering crossovers were unaffected. We have also identified a physical interaction between UBC-9 and DNA repair protein MRE-11 and null mutants exhibited similar phenotypes at germline mitotic nuclei and were synthetically sick. These phenotypes and genetic interactions were specific to MRE-11 null mutants as opposed to RAD-50 or resection-defective MRE-11 We propose that the SUMOylation pathway acts redundantly with MRE-11, and in this process MRE-11 likely plays a structural role.
减数分裂是一个高度调控的过程,部分原因是需要断裂和修复 DNA,作为减数分裂程序的一部分。翻译后修饰在减数分裂事件中广泛用于调节蛋白质复合物形成、检查点激活和蛋白质衰减等步骤。在本文中,我们研究了作为一种翻译后修饰的 SUMO(小泛素样修饰物)途径的必需组成部分的蛋白质如何影响生殖细胞。我们表明,E2 连接酶 UBC-9 和 SUMO 的同源物 SMO-1 在整个前期 I 中定位于生殖细胞核。对 和 的突变分析表明,生殖细胞中存在更多的重组中间体,这些中间体起源于有丝分裂分裂。SUMOylation 突变体也表现出晚期减数分裂缺陷,包括卵母细胞二价体的重构和有丝分裂卵母细胞的缺陷。在 杂合子中观察到非干扰交叉的频率增加,尽管干扰交叉不受影响。我们还鉴定了 UBC-9 和 DNA 修复蛋白 MRE-11 之间的物理相互作用,并且 和 缺失突变体在生殖细胞有丝分裂核中表现出相似的表型,并表现出合成性疾病。这些表型和遗传相互作用是 MRE-11 缺失突变体特有的,而不是 RAD-50 或切除缺陷型 MRE-11 缺失突变体。我们提出 SUMOylation 途径与 MRE-11 冗余作用,在此过程中,MRE-11 可能发挥结构作用。