Singapore Institute of Technology, Singapore.
Bioinformatics Institute, A*STAR, Singapore.
PLoS Comput Biol. 2018 Mar 5;14(3):e1005966. doi: 10.1371/journal.pcbi.1005966. eCollection 2018 Mar.
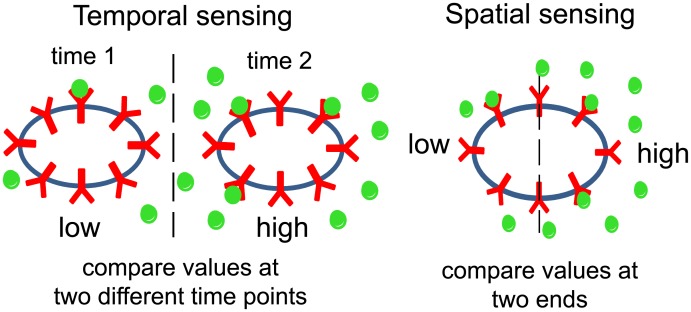
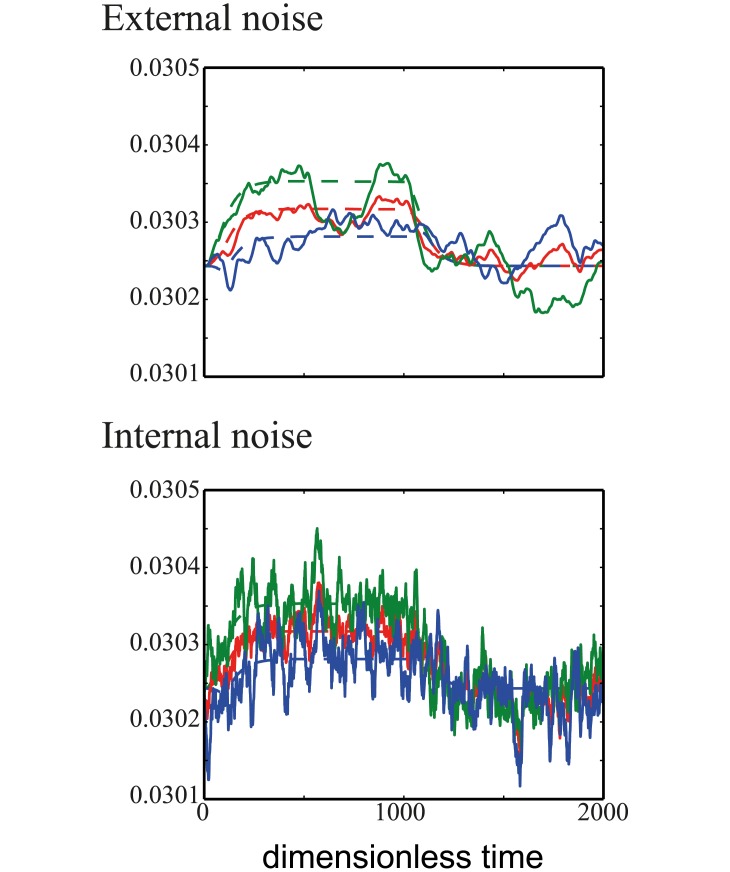
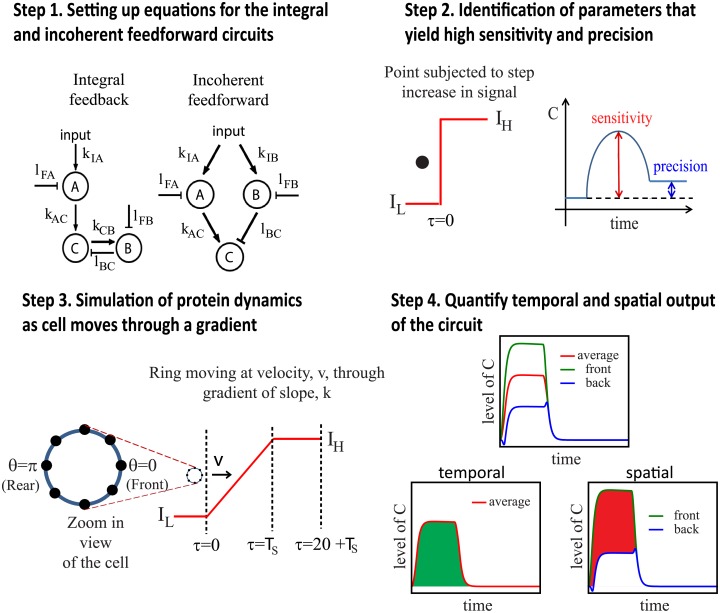
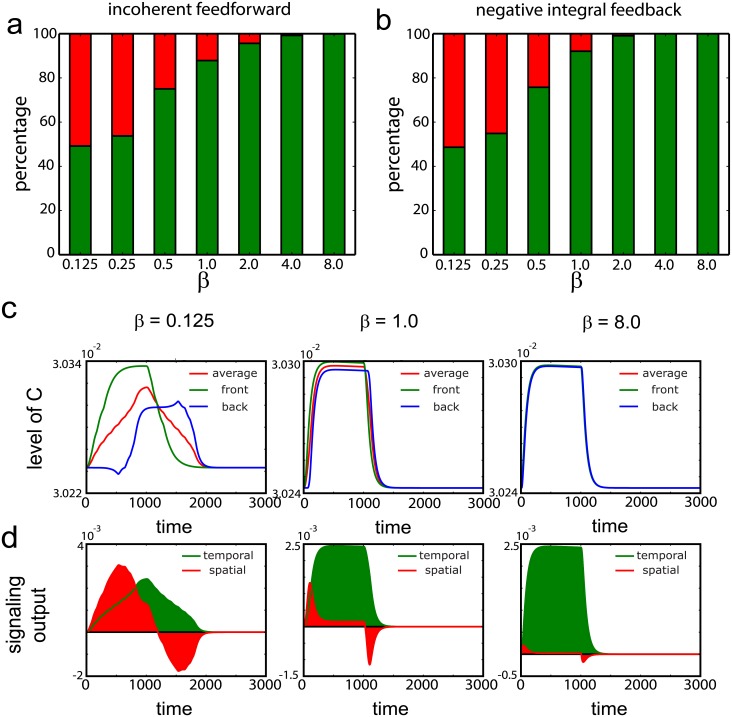
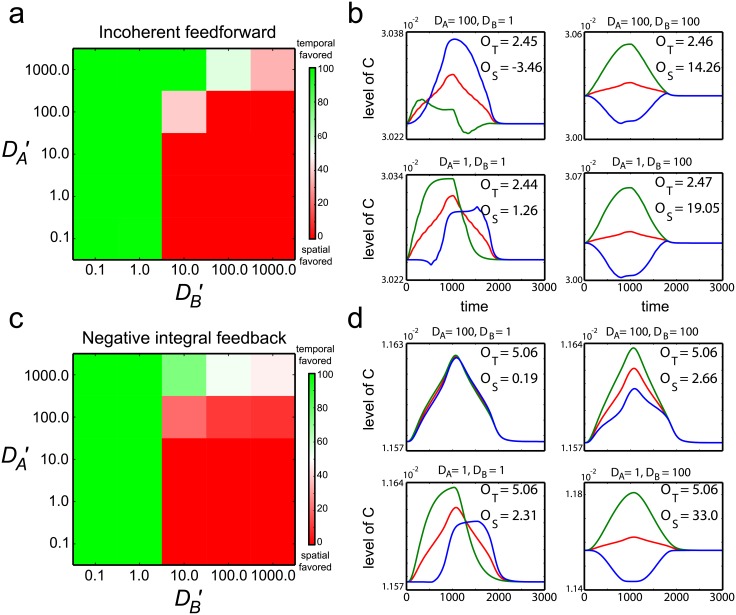
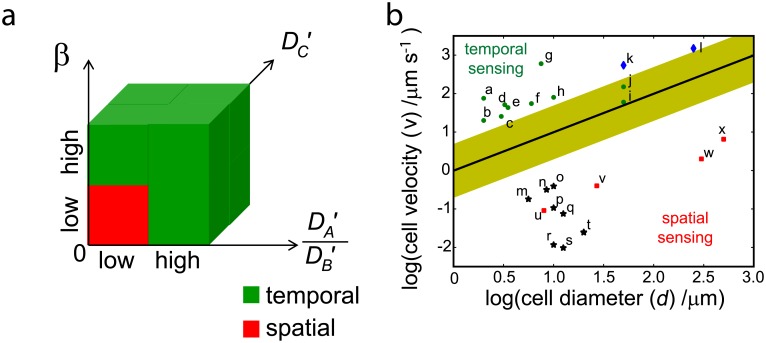
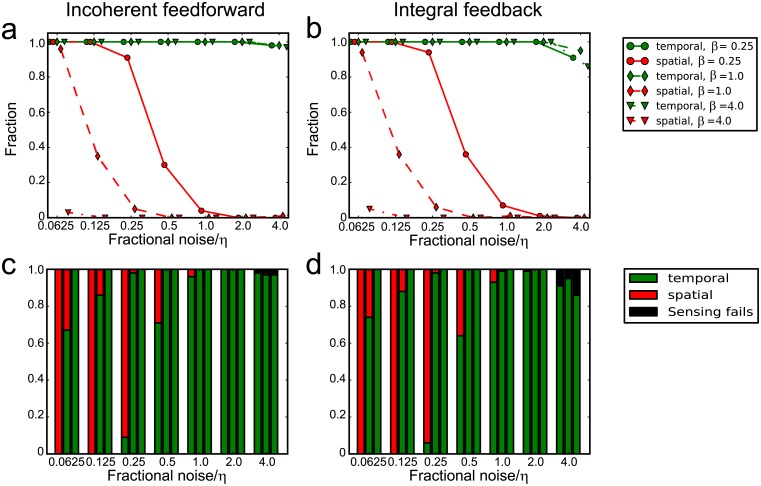
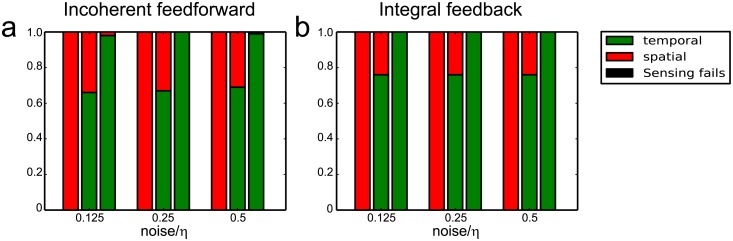
Cell size is thought to play an important role in choosing between temporal and spatial sensing in chemotaxis. Large cells are thought to use spatial sensing due to large chemical difference at its ends whereas small cells are incapable of spatial sensing due to rapid homogenization of proteins within the cell. However, small cells have been found to polarize and large cells like sperm cells undergo temporal sensing. Thus, it remains an open question what exactly governs spatial versus temporal sensing. Here, we identify the factors that determines sensing choices through mathematical modeling of chemotactic circuits. Comprehensive computational search of three-node signaling circuits has identified the negative integral feedback (NFB) and incoherent feedforward (IFF) circuits as capable of adaptation, an important property for chemotaxis. Cells are modeled as one-dimensional circular system consisting of diffusible activator, inactivator and output proteins, traveling across a chemical gradient. From our simulations, we find that sensing outcomes are similar for NFB or IFF circuits. Rather than cell size, the relevant parameters are the 1) ratio of cell speed to the product of cell diameter and rate of signaling, 2) diffusivity of the output protein and 3) ratio of the diffusivities of the activator to inactivator protein. Spatial sensing is favored when all three parameters are low. This corresponds to a cell moving slower than the time it takes for signaling to propagate across the cell diameter, has an output protein that is polarizable and has a local-excitation global-inhibition system to amplify the chemical gradient. Temporal sensing is favored otherwise. We also find that temporal sensing is more robust to noise. By performing extensive literature search, we find that our prediction agrees with observation in a wide range of species and cell types ranging from E. coli to human Fibroblast cells and propose that our result is universally applicable.
细胞大小被认为在选择趋化性中的时空感应中起着重要作用。大细胞由于其两端的化学差异较大,因此被认为使用空间感应,而小细胞由于细胞内蛋白质的快速均匀化而无法进行空间感应。然而,已经发现小细胞能够极化,而大细胞如精子细胞则进行时间感应。因此,究竟是什么决定了空间感应和时间感应,这仍然是一个悬而未决的问题。在这里,我们通过对趋化性电路的数学建模来确定决定感应选择的因素。对三节点信号电路的全面计算搜索已经确定了负积分反馈(NFB)和非相干前馈(IFF)电路具有适应性,这是趋化性的一个重要特性。细胞被建模为一维圆形系统,由可扩散的激活剂、失活剂和输出蛋白组成,在化学梯度中移动。从我们的模拟中,我们发现 NFB 或 IFF 电路的感应结果相似。而不是细胞大小,相关参数是 1)细胞速度与细胞直径和信号传递速率的乘积之比,2)输出蛋白的扩散率和 3)激活剂和失活剂蛋白扩散率之比。当所有三个参数都较低时,空间感应是有利的。这对应于细胞移动速度比信号传播穿过细胞直径所需的时间慢,具有可极化的输出蛋白,并且具有局部兴奋全局抑制系统来放大化学梯度。否则,时间感应是有利的。我们还发现,时间感应对噪声的鲁棒性更强。通过进行广泛的文献搜索,我们发现我们的预测与从大肠杆菌到人类成纤维细胞的广泛物种和细胞类型的观察结果一致,并提出我们的结果普遍适用。