CNRS, Univ. Lille, Univ. Littoral Côte d'Opale, UMR 8187, LOG, Laboratoire d'Océanologie et de Géosciences, Wimereux, France.
Sir Alister Hardy Foundation for Ocean Science, The Laboratory, Citadel Hill, Plymouth, United Kingdom.
PLoS One. 2018 Mar 22;13(3):e0194006. doi: 10.1371/journal.pone.0194006. eCollection 2018.
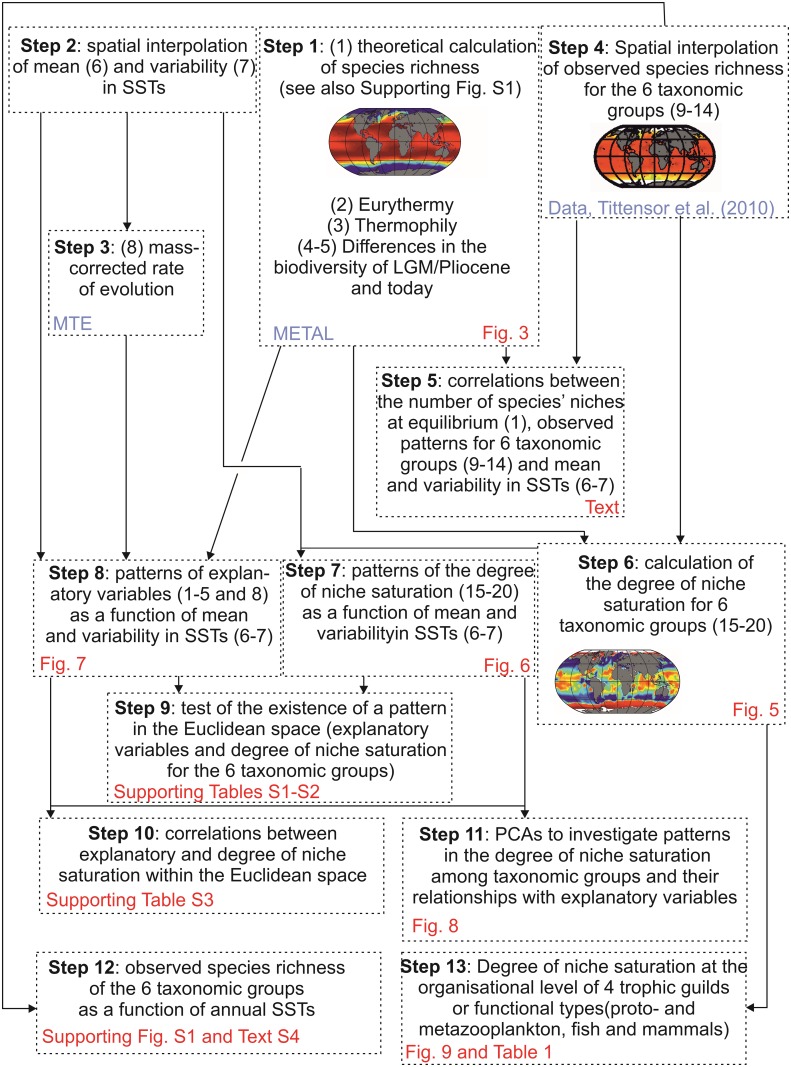
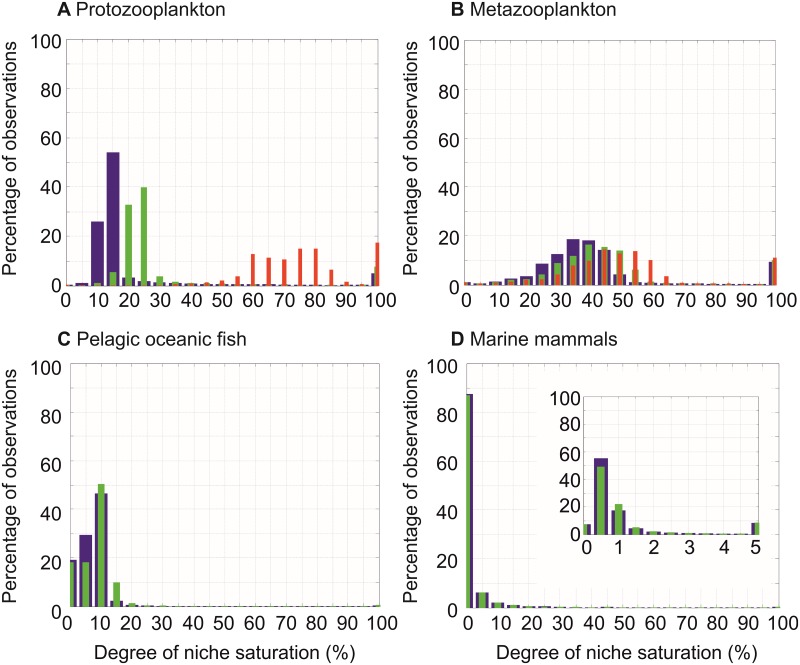
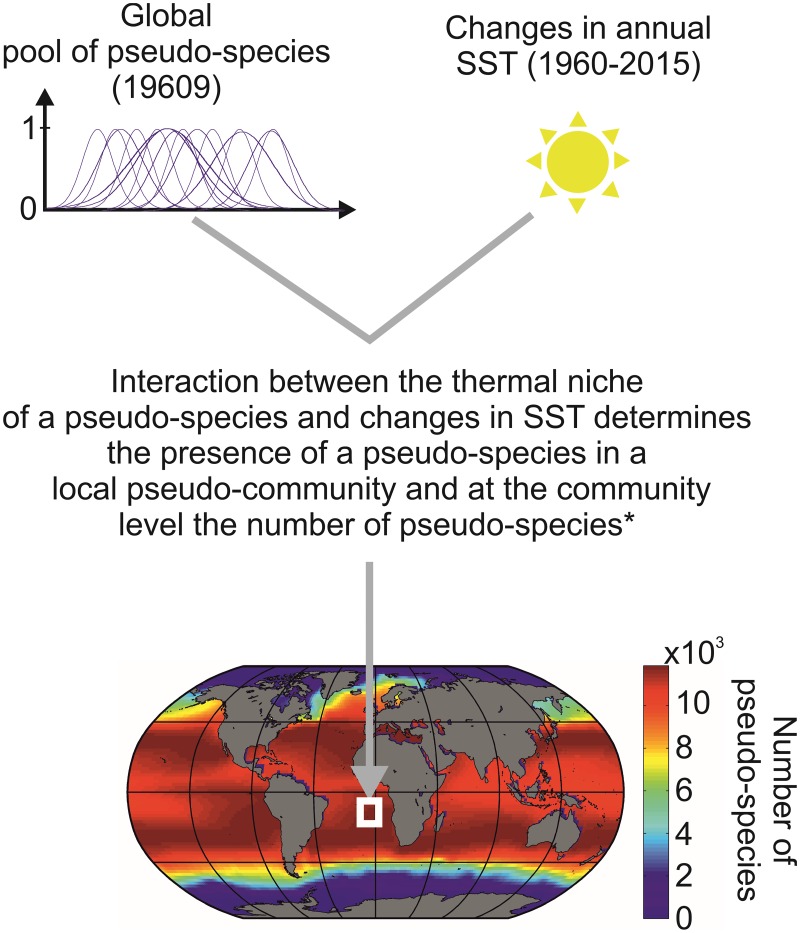
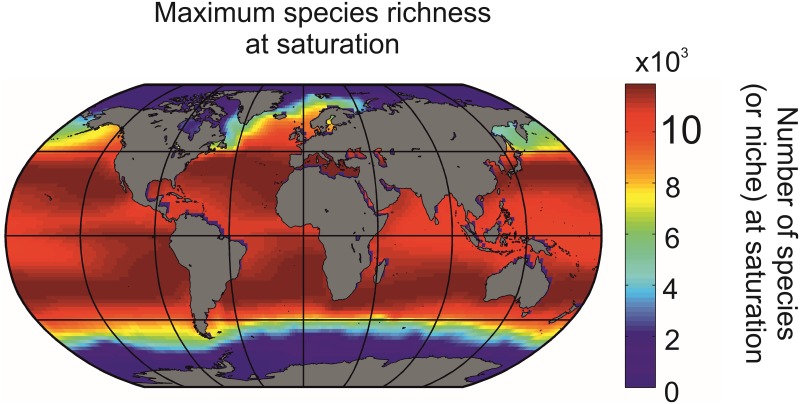
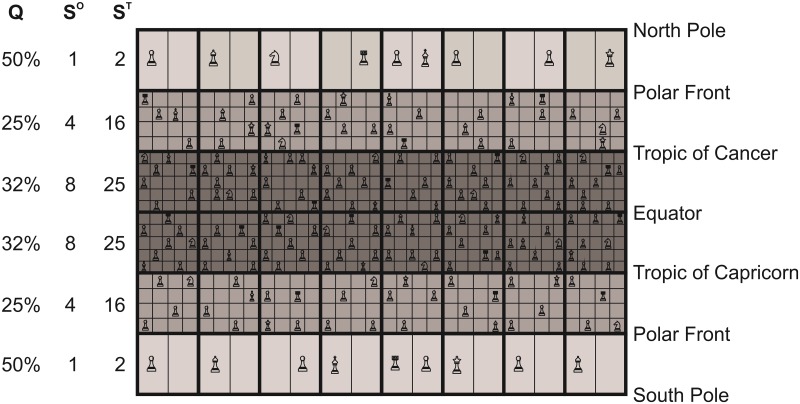
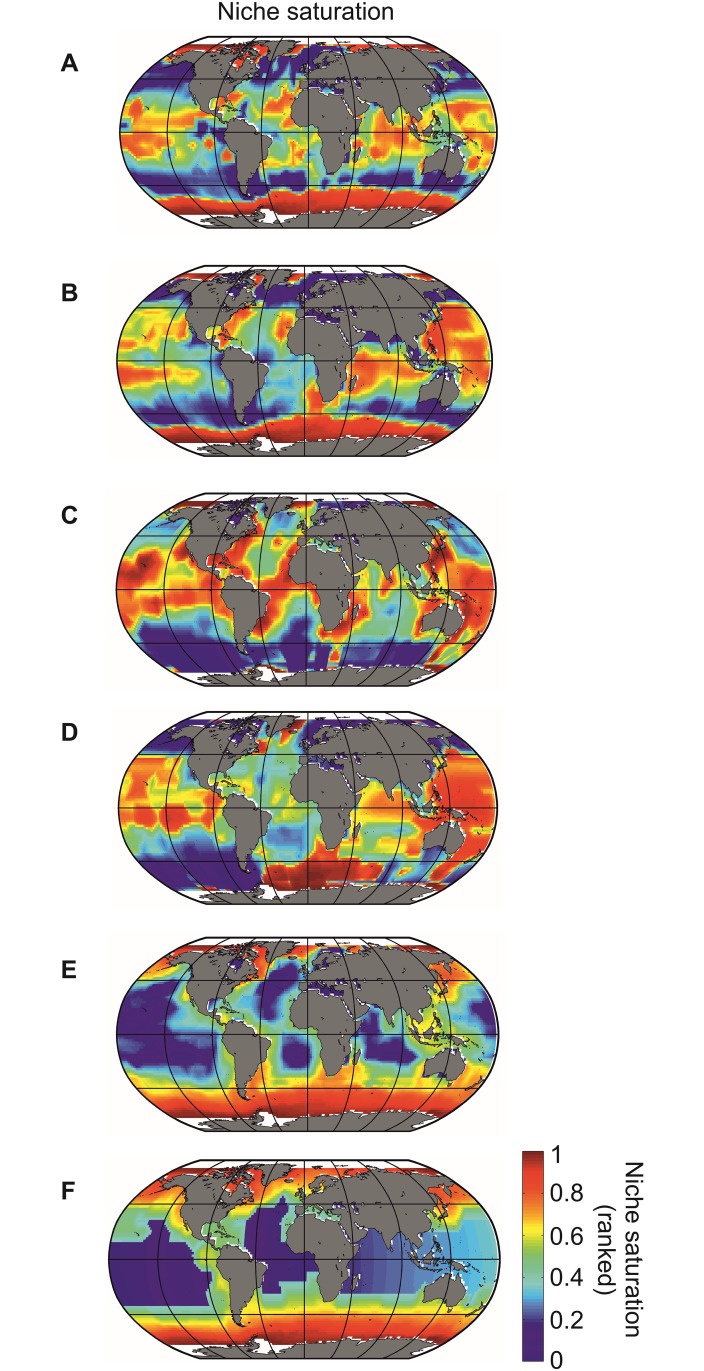
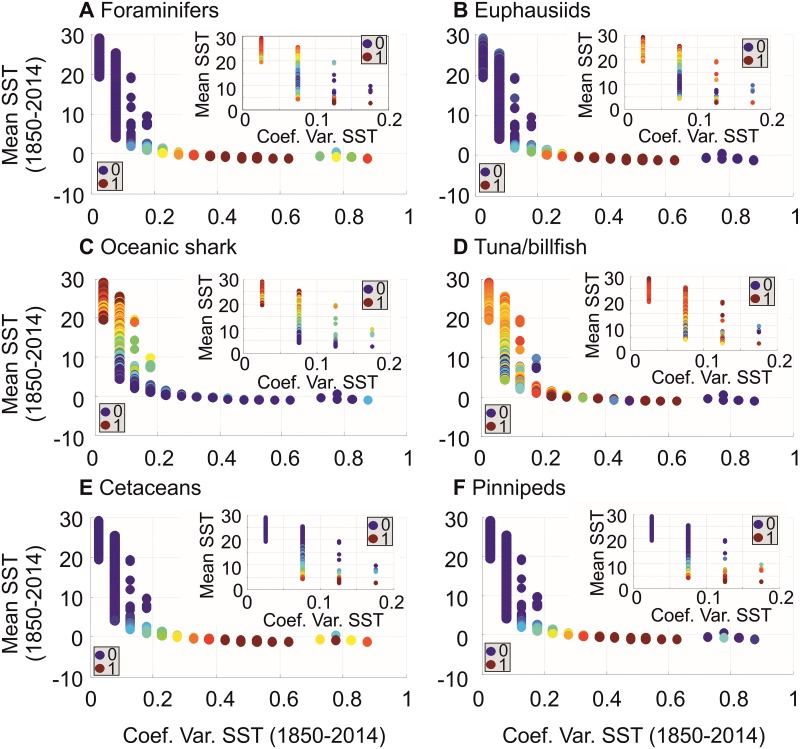
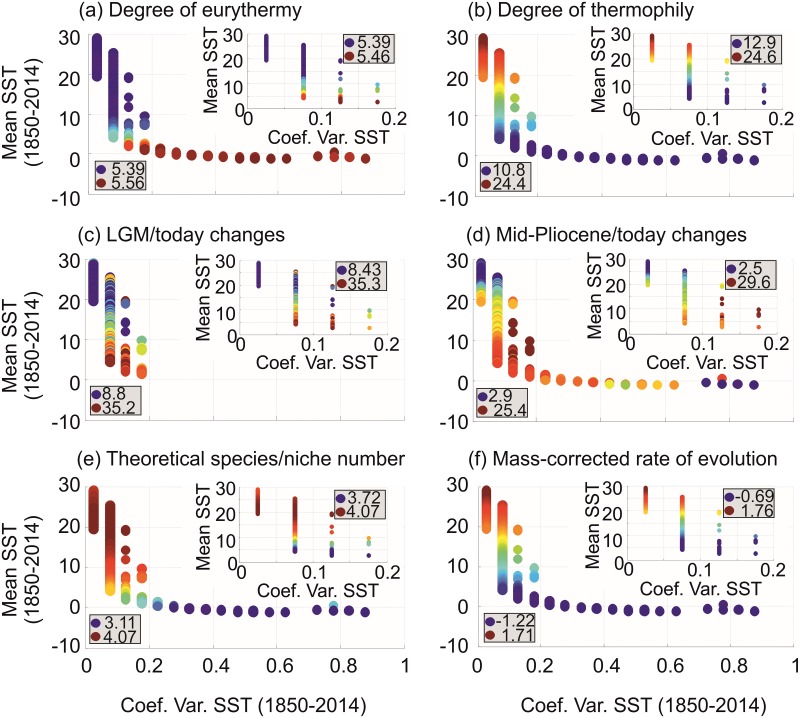
Species richness is greater in places where the number of potential niches is high. Consequently, the niche may be fundamental for understanding the arrangement of life and especially, the establishment and maintenance of the well-known Latitudinal Biodiversity Gradient (LBG). However, not all potential niches may be occupied fully in a habitat, as measured by niche vacancy/saturation. Here, we theoretically reconstruct oceanic biodiversity and analyse modeled and observed data together to examine patterns in niche saturation (i.e. the ratio between observed and theoretical biodiversity of a given taxon) for several taxonomic groups. Our results led us to hypothesize that the arrangement of marine life is constrained by the distribution of the maximal number of species' niches available, which represents a fundamental mathematical limit to the number of species that can co-exist locally. We liken this arrangement to a type of chessboard where each square on the board is a geographic area, itself comprising a distinct number of sub-squares (species' niches). Each sub-square on the chessboard can accept a unique species of a given ecological guild, whose occurrence is determined by speciation/extinction. Because of the interaction between the thermal niche and changes in temperature, our study shows that the chessboard has more sub-squares at mid-latitudes and we suggest that many clades should exhibit a LBG because their probability of emergence should be higher in the tropics where more niches are available. Our work reveals that each taxonomic group has its own unique chessboard and that global niche saturation increases when organismal complexity decreases. As a result, the mathematical influence of the chessboard is likely to be more prominent for taxonomic groups with low (e.g. plankton) than great (e.g. mammals) biocomplexity. Our study therefore reveals the complex interplay between a fundamental mathematical constraint on biodiversity resulting from the interaction between the species' ecological niche and fluctuations in the environmental regime (here, temperature), which has a predictable component and a stochastic-like biological influence (diversification rates, origination and clade age) that may alter or blur the former.
物种丰富度在潜在生态位数量较高的地方更高。因此,生态位可能是理解生命排列的基础,尤其是著名的纬度生物多样性梯度(LBG)的建立和维持的基础。然而,在一个栖息地中,并非所有潜在的生态位都被完全占据,这可以通过生态位空缺/饱和来衡量。在这里,我们从理论上重建了海洋生物多样性,并对模型和观测数据进行了综合分析,以研究几个分类群的生态位饱和模式(即给定分类群的观测生物多样性与理论生物多样性之比)。我们的结果使我们假设,海洋生物的排列受到可用最大物种生态位数量的分布限制,这代表了本地共存的物种数量的基本数学限制。我们将这种排列比作一种棋盘,棋盘上的每个方格代表一个地理区域,本身包含一定数量的小方格(物种生态位)。棋盘上的每个小方格可以容纳一个给定生态类群的独特物种,其出现是由物种形成/灭绝决定的。由于热生态位和温度变化之间的相互作用,我们的研究表明,棋盘在中纬度地区有更多的小方格,我们认为许多进化枝应该表现出 LBG,因为它们在有更多生态位的热带地区出现的概率更高。我们的工作表明,每个分类群都有自己独特的棋盘,并且当生物体复杂性降低时,全球生态位饱和度会增加。因此,棋盘的数学影响对于生物复杂性低的分类群(例如浮游生物)可能比生物复杂性高的分类群(例如哺乳动物)更为显著。因此,我们的研究揭示了生物多样性的基本数学限制与物种生态位和环境变化(此处为温度)之间相互作用的复杂相互作用,这种相互作用具有可预测的成分和类似随机的生物学影响(多样化率、起源和进化枝年龄),这些影响可能会改变或模糊前者。