Center for Genomics and Computational Biology, Department of Biomedical Engineering, Duke University, Durham, United States.
Sibley School of Mechanical and Aerospace Engineering, Cornell University, Ithaca, United States.
Elife. 2018 Apr 9;7:e35710. doi: 10.7554/eLife.35710.
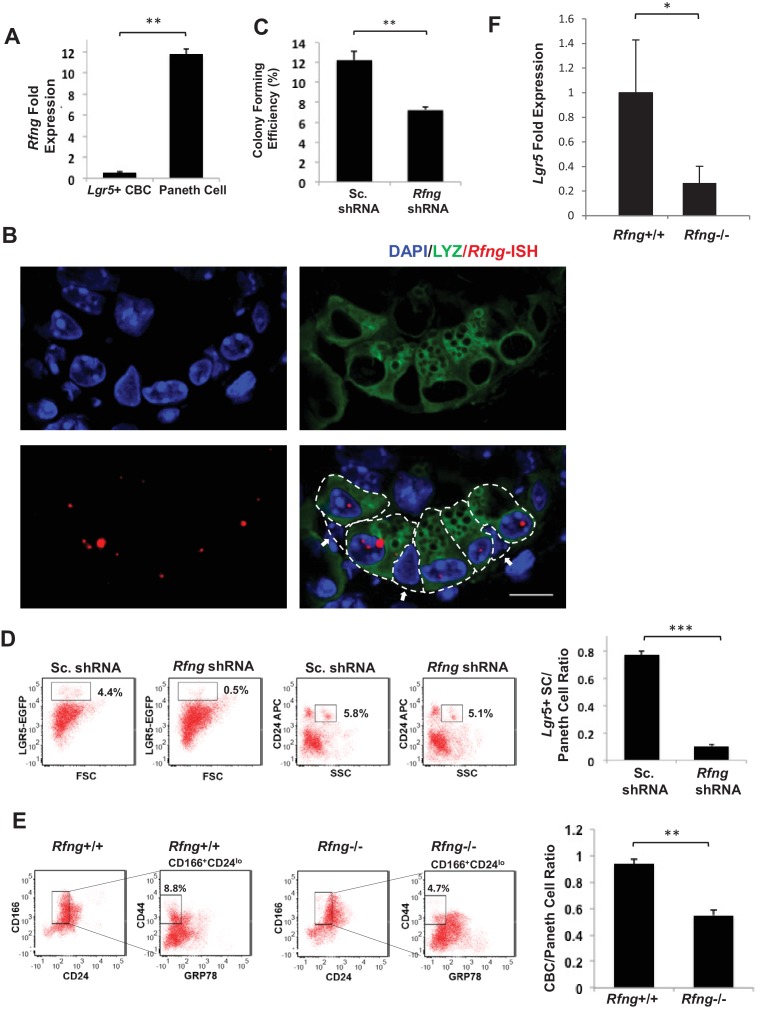
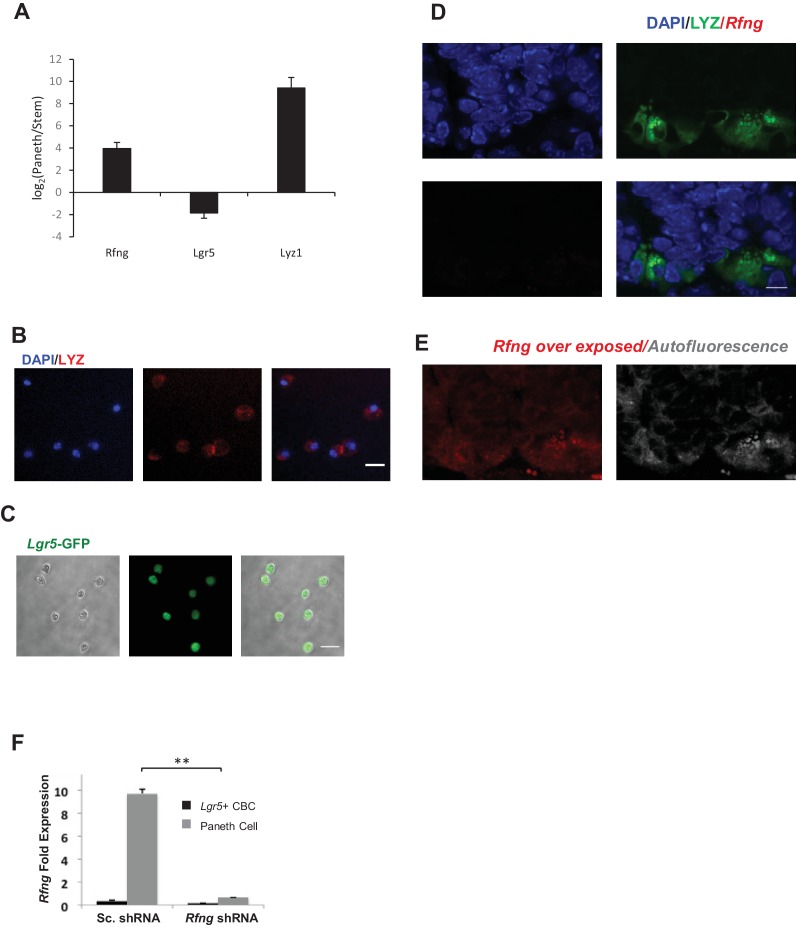
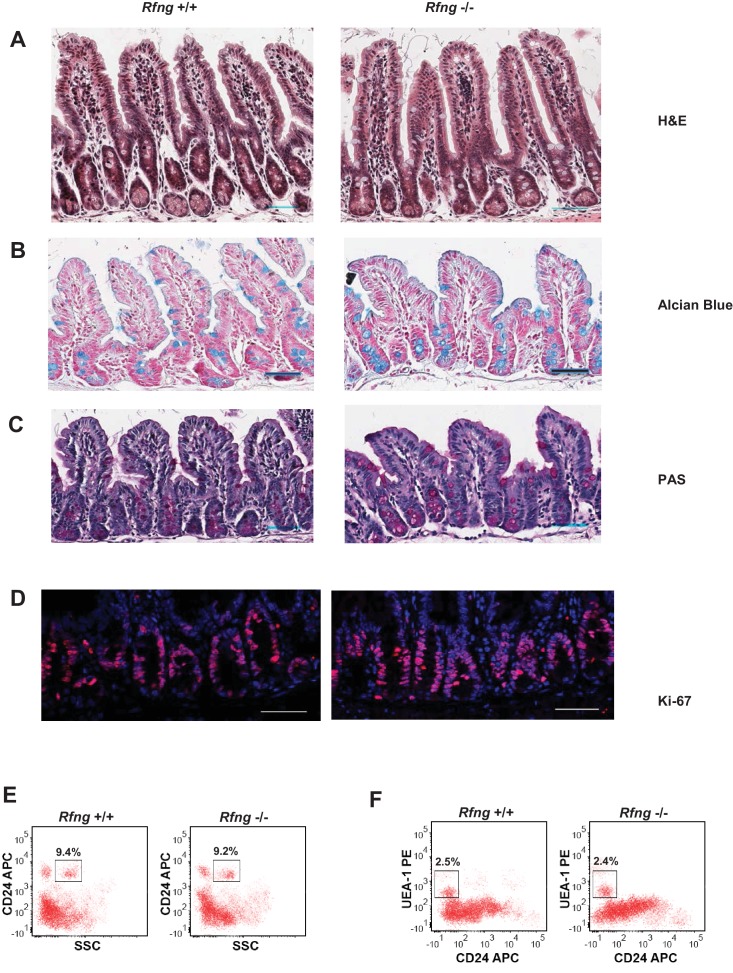
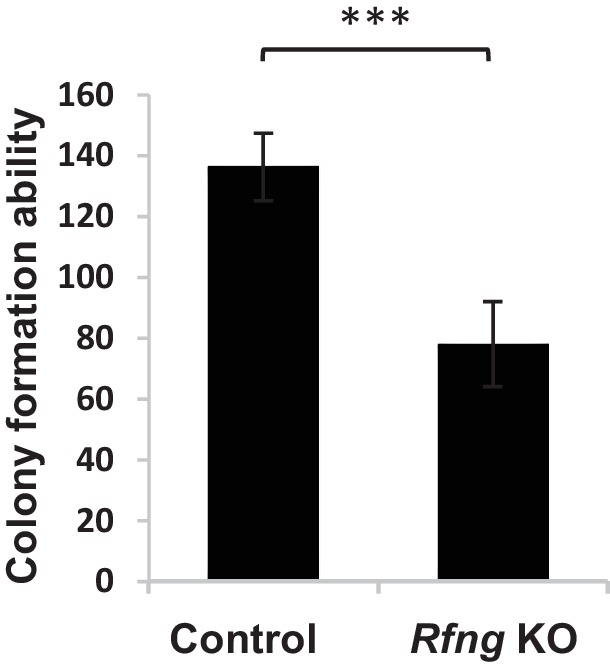
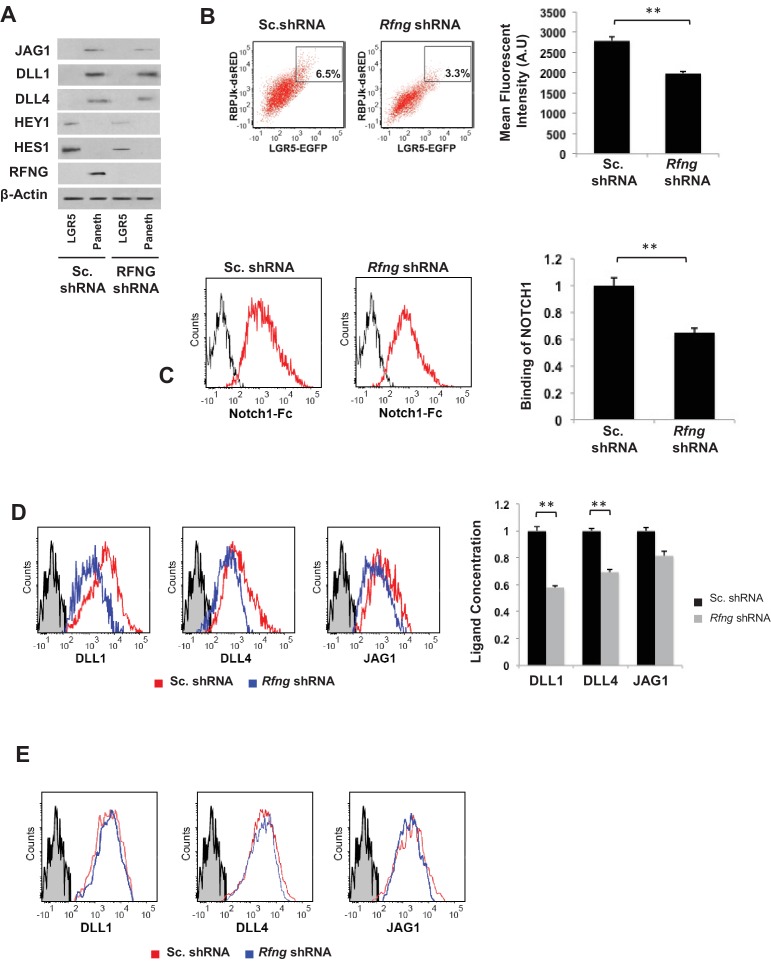
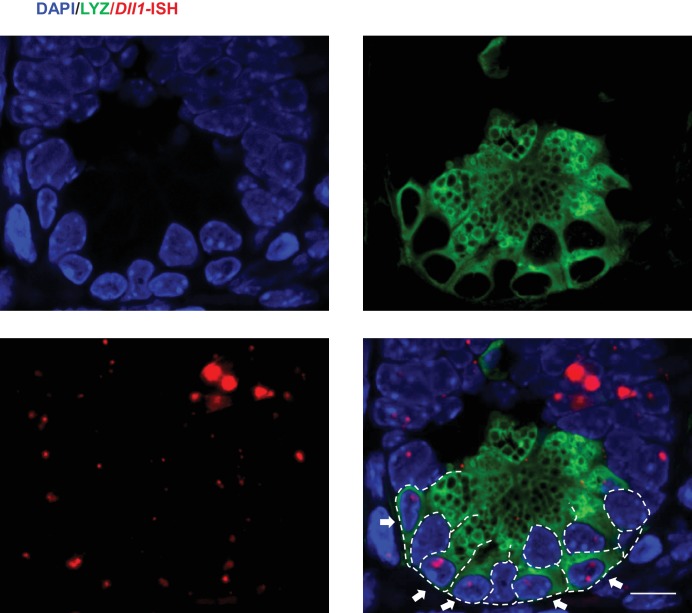
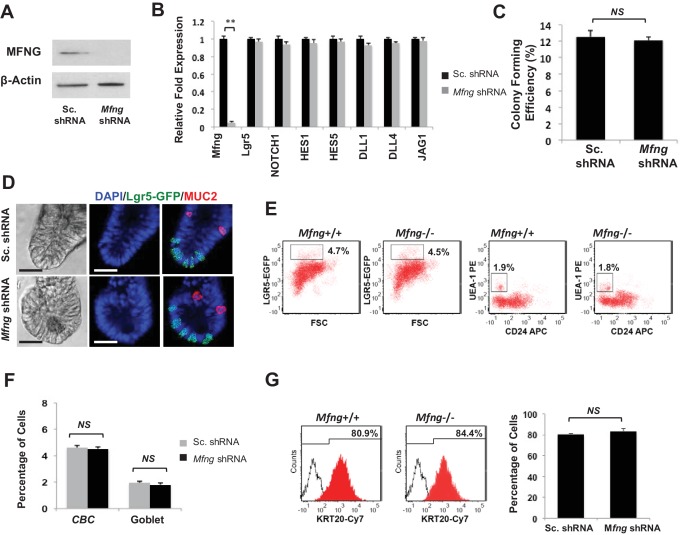
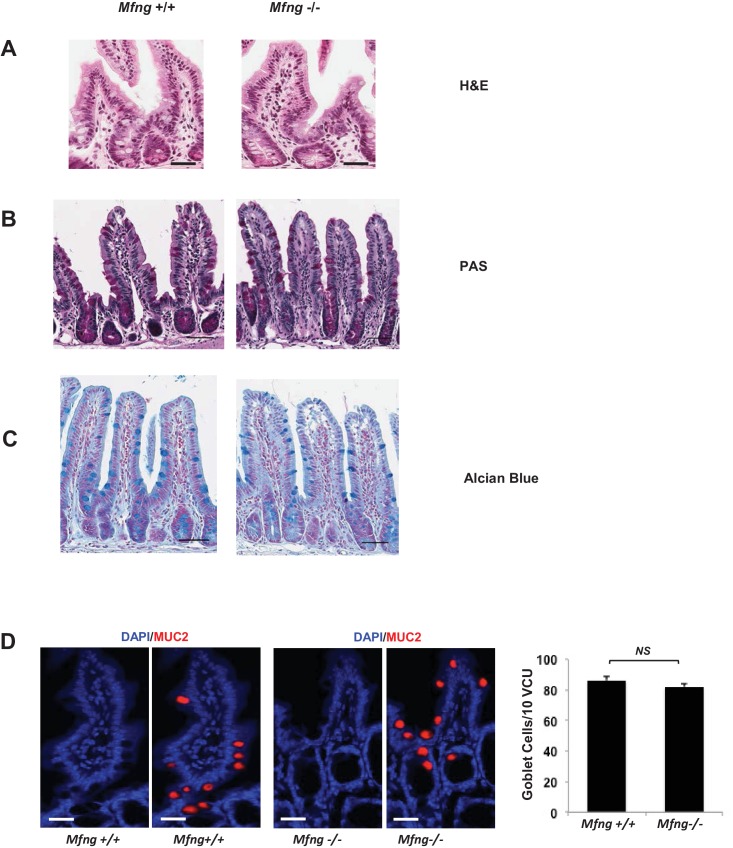
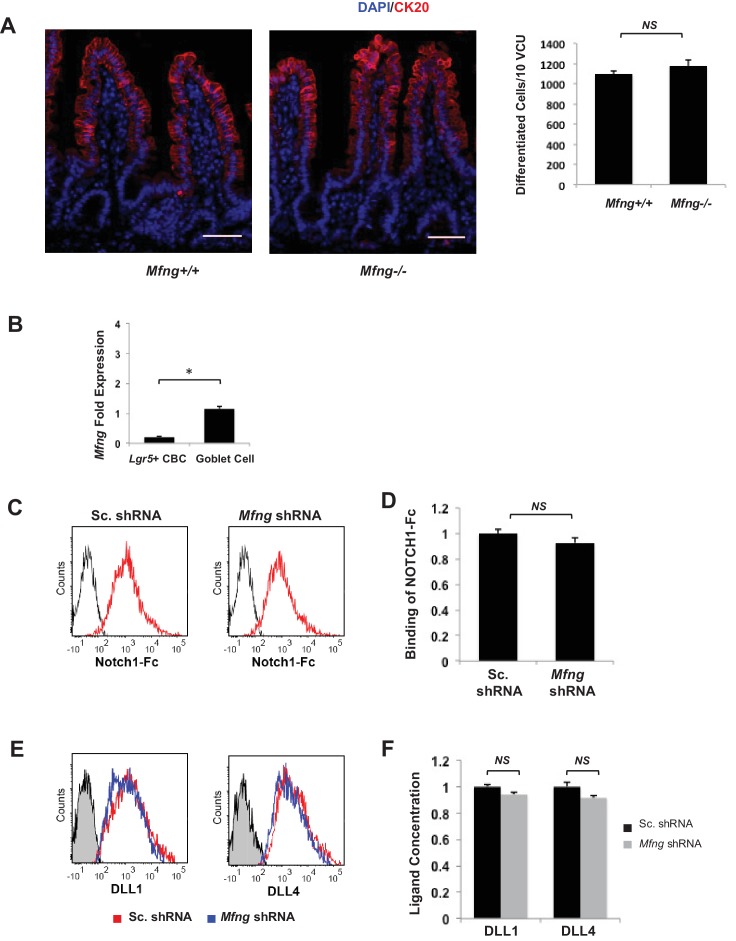
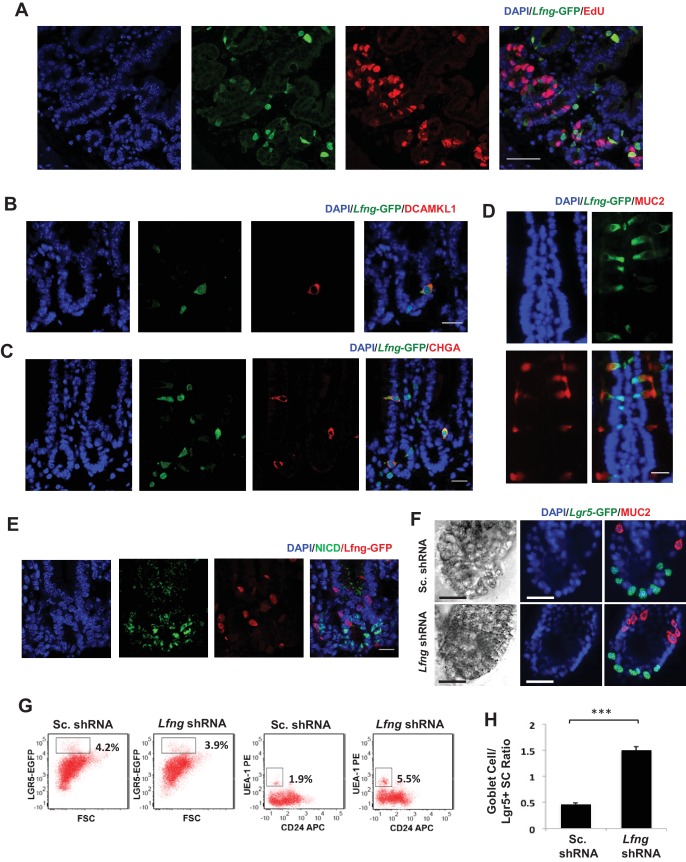
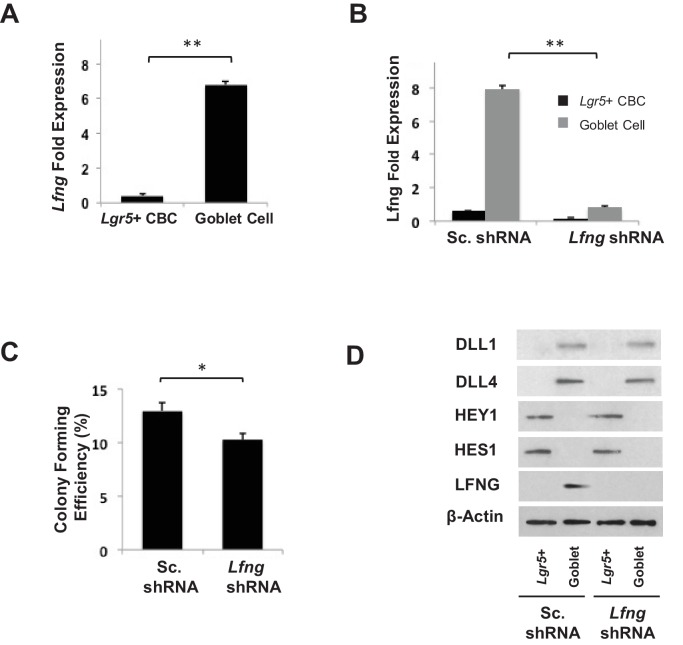
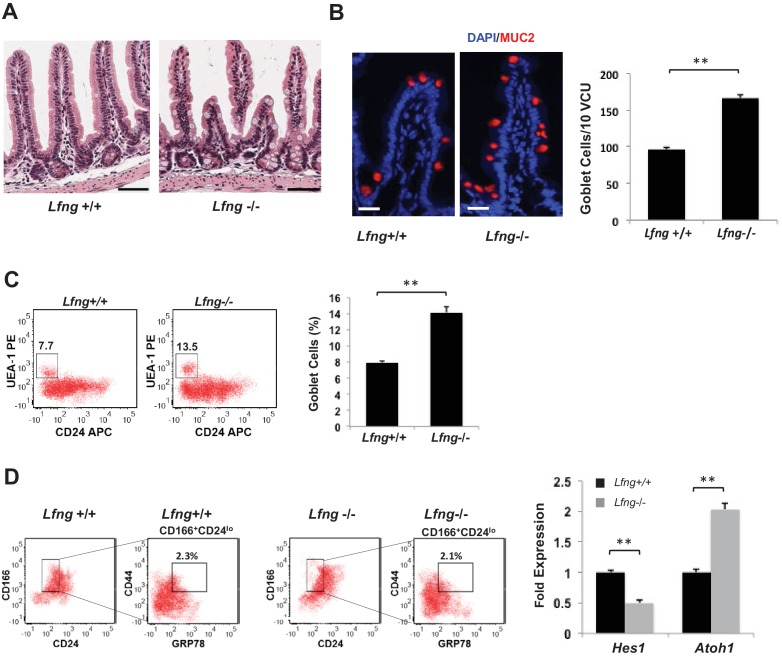
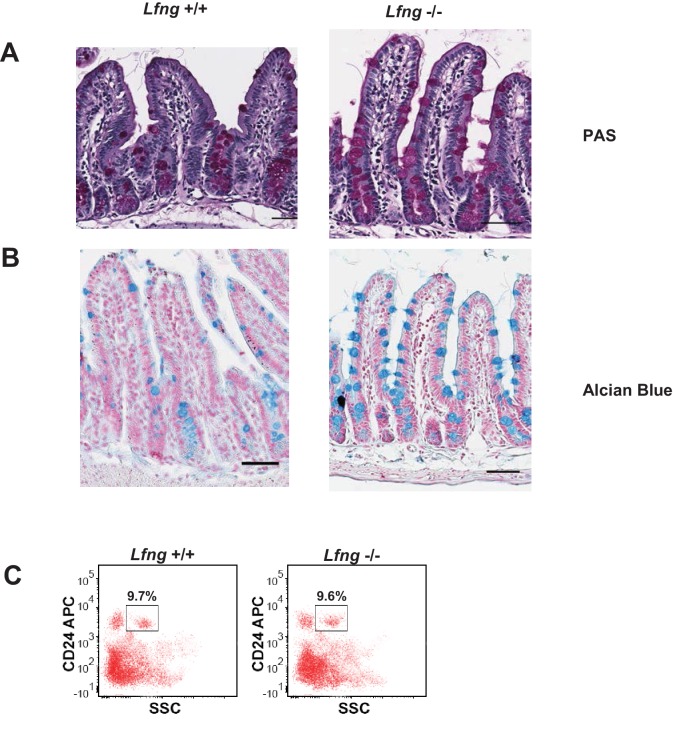
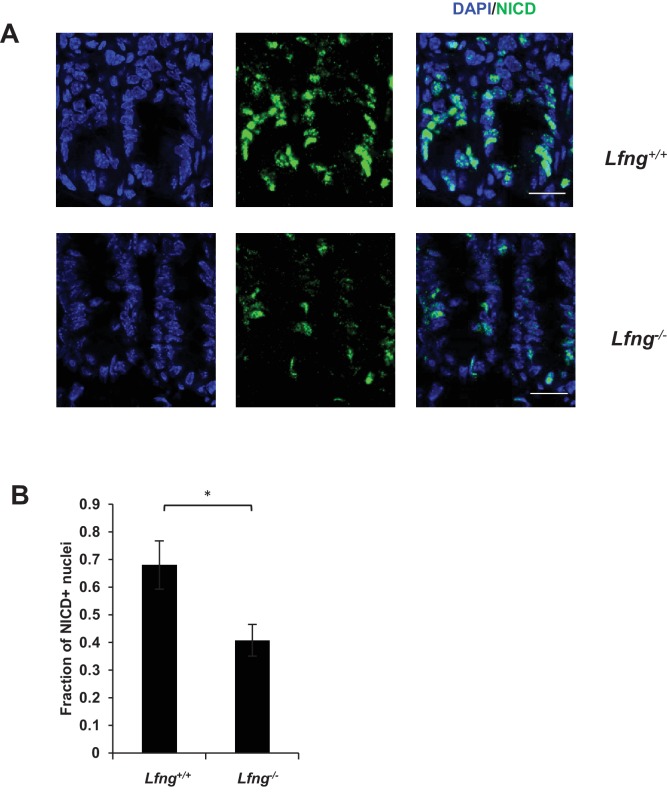
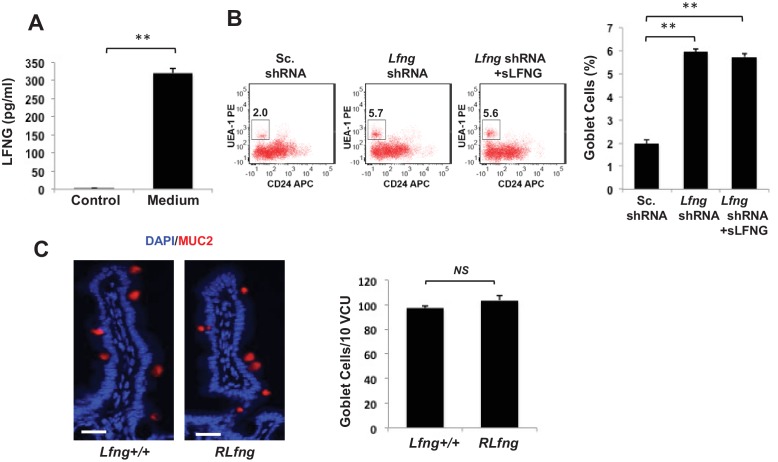
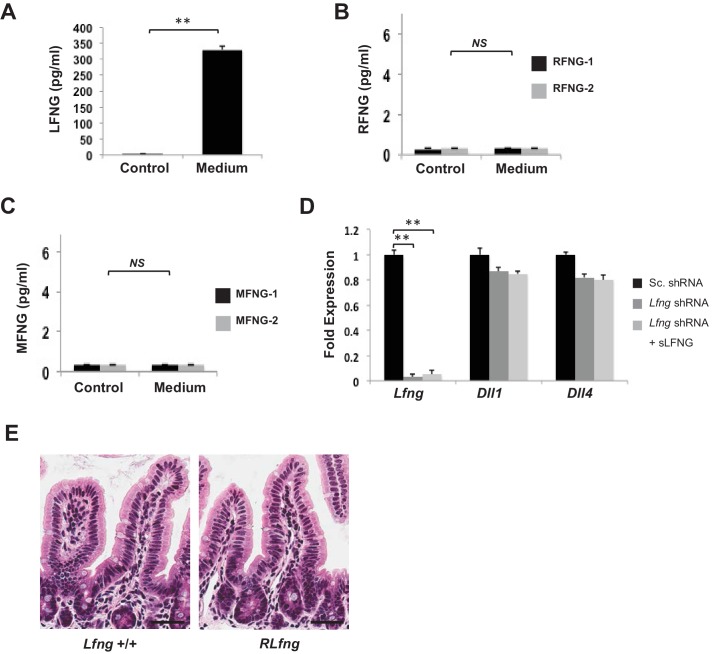
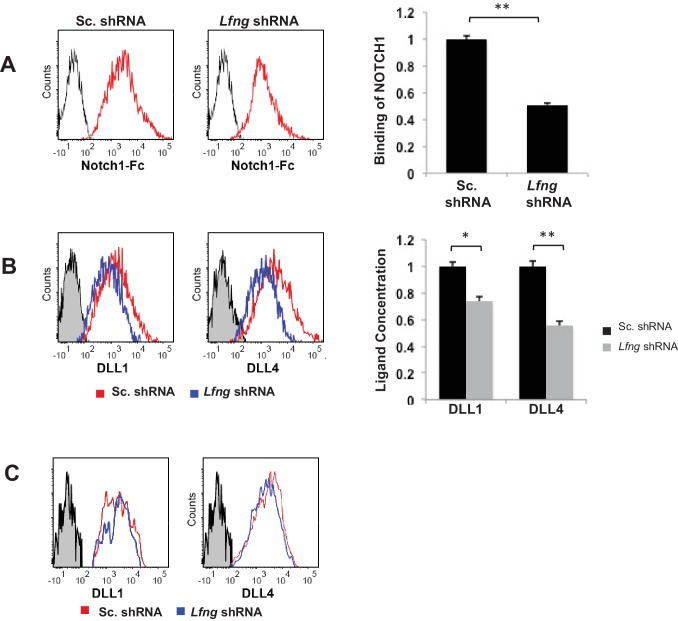
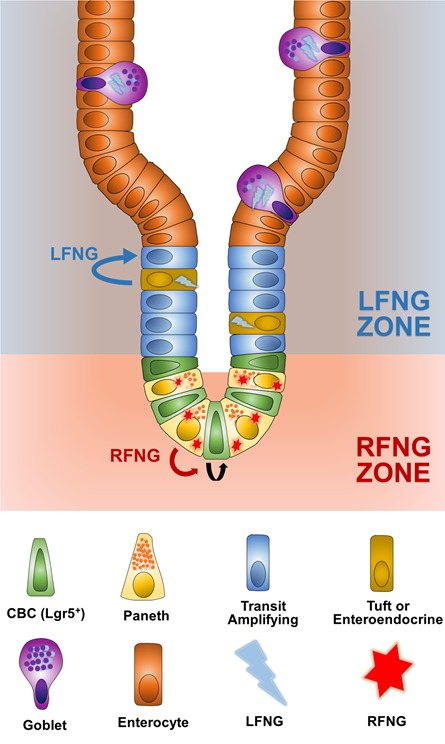
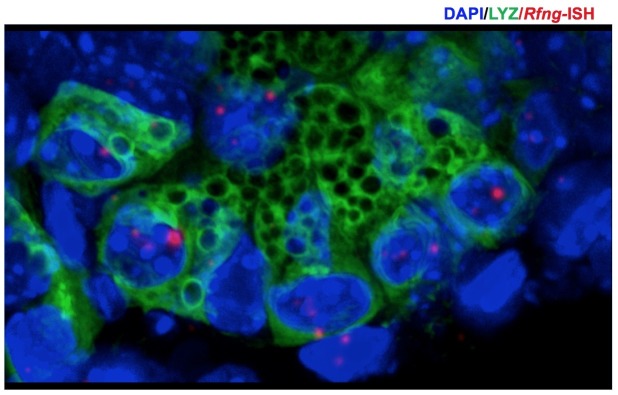
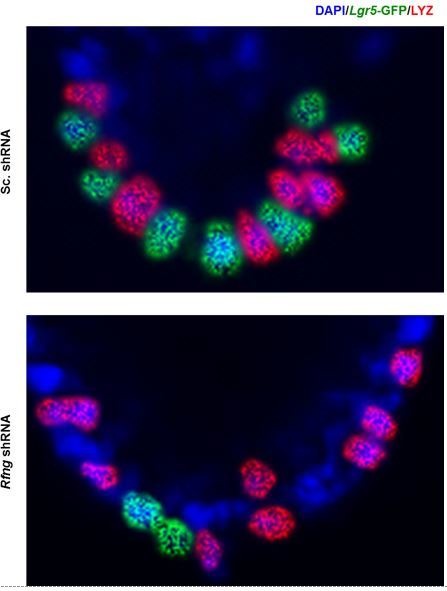
Notch signalling maintains stem cell regeneration at the mouse intestinal crypt base and balances the absorptive and secretory lineages in the upper crypt and villus. Here we report the role of Fringe family of glycosyltransferases in modulating Notch activity in the two compartments. At the crypt base, RFNG is enriched in the Paneth cells and increases cell surface expression of DLL1 and DLL4. This promotes Notch activity in the neighbouring + stem cells assisting their self-renewal. Expressed by various secretory cells in the upper crypt and villus, LFNG promotes DLL surface expression and suppresses the secretory lineage . Hence, in the intestinal epithelium, Fringes are present in the ligand-presenting 'sender' secretory cells and promote Notch activity in the neighbouring 'receiver' cells. Fringes thereby provide for targeted modulation of Notch activity and thus the cell fate in the stem cell zone, or the upper crypt and villus.
Notch 信号通路在维持肠道隐窝底部干细胞的再生中起着关键作用,并平衡了隐窝和绒毛中吸收细胞和分泌细胞的谱系分化。在此,我们报告了 Fringe 家族糖基转移酶在调节两个隔室中的 Notch 活性中的作用。在隐窝底部,RFNG 富含于 Paneth 细胞中,并增加 DLL1 和 DLL4 的细胞表面表达。这促进了相邻的 +干细胞中的 Notch 活性,有助于它们的自我更新。LFNG 由隐窝和绒毛中的各种分泌细胞表达,促进 DLL 的表面表达并抑制分泌谱系。因此,在肠道上皮中,Fringes 存在于配体呈递的“发送”分泌细胞中,并促进相邻“接收”细胞中的 Notch 活性。Fringes 从而为 Notch 活性和干细胞区或隐窝和绒毛中的细胞命运提供了靶向调节。