Wang Zhixing, An Ning, Xu Wenqiang, Zhang Weixin, Meng Xiangfeng, Chen Guanjun, Liu Weifeng
State Key Laboratory of Microbial Technology, School of Life Science, Shandong University, No.27 Shanda South Road, Jinan, 250100 Shandong People's Republic of China.
Biotechnol Biofuels. 2018 Apr 4;11:97. doi: 10.1186/s13068-018-1098-8. eCollection 2018.
holds a high capacity for protein secretion and represents the most important cellulase producer in industry. However, the external signal sensing and intracellular signal transduction during cellulose induction remain unclear. As one of the most pervasive signal transduction pathways in all eukaryotic species, the mitogen-activated protein kinase (MAPK) pathway and its upstream sensing and signaling components are involved in various physiological processes including stress and nutrient sensing. Particularly, the Hog1-type MAPK Tmk3 has been reported to be involved in the cellulase production in .
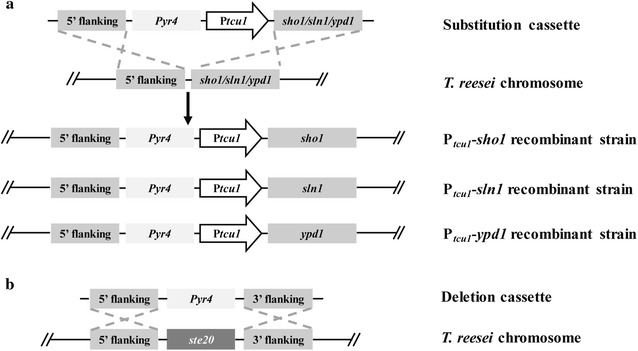
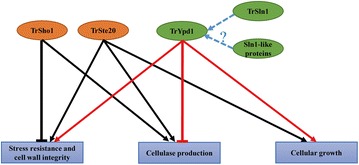
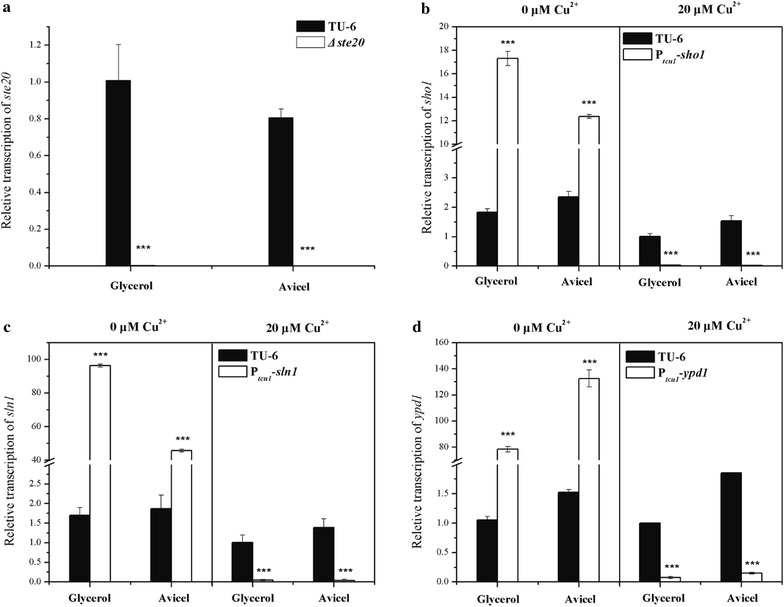
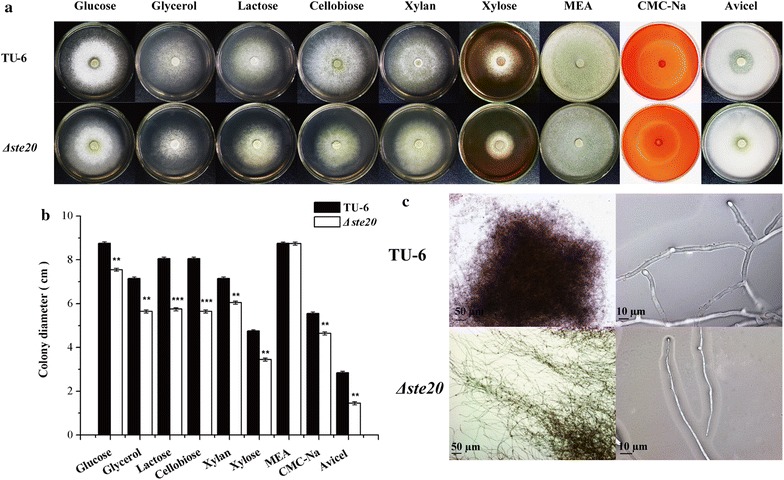
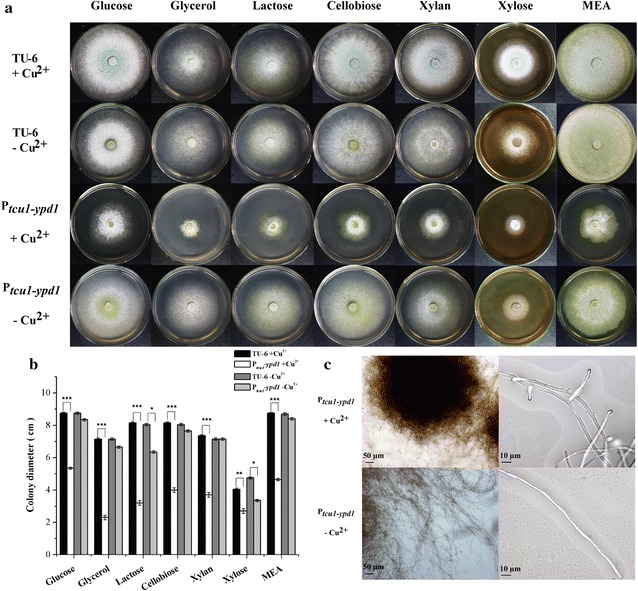
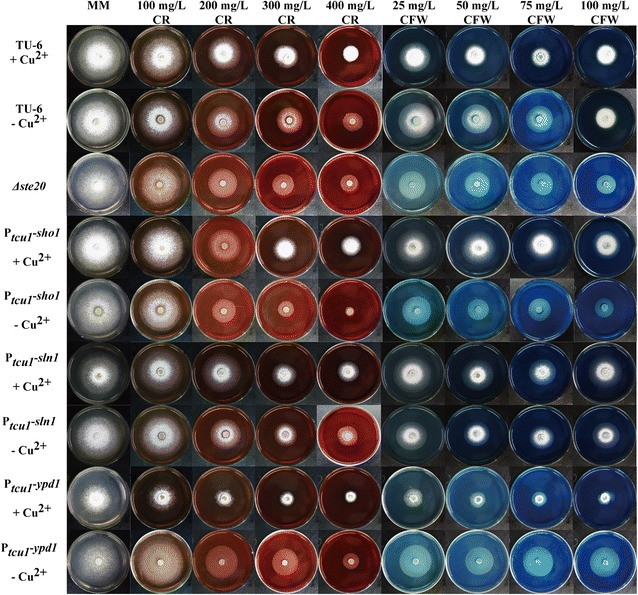
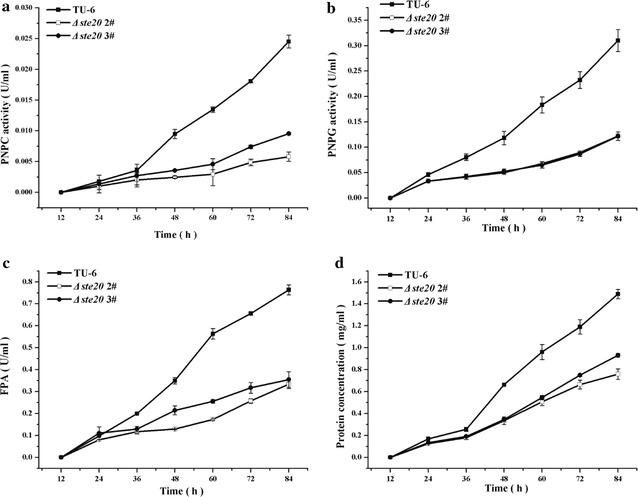
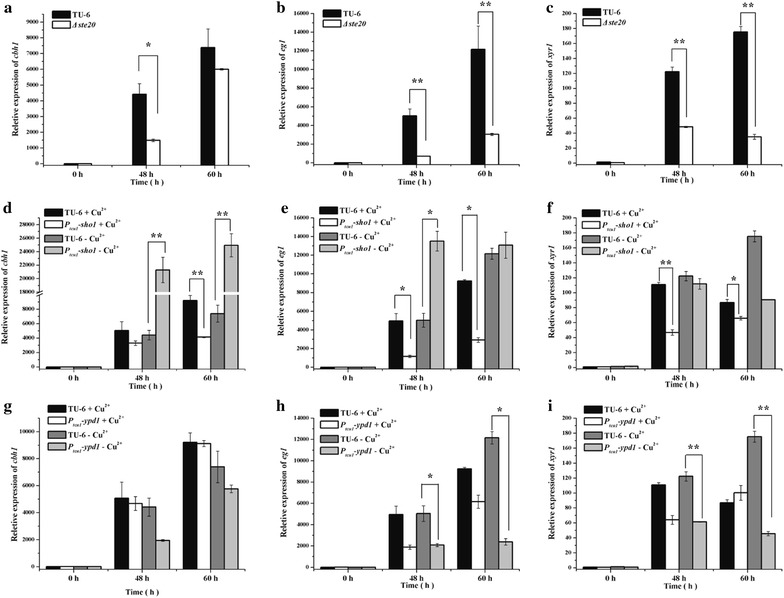
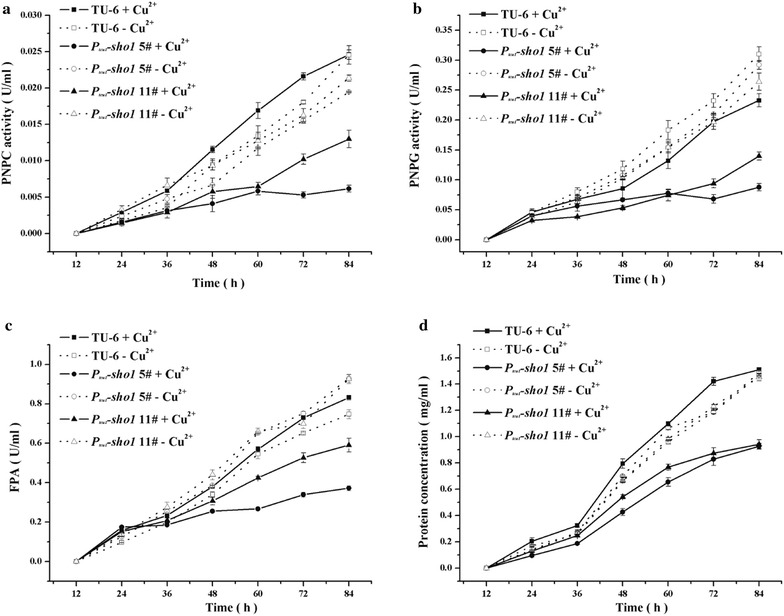
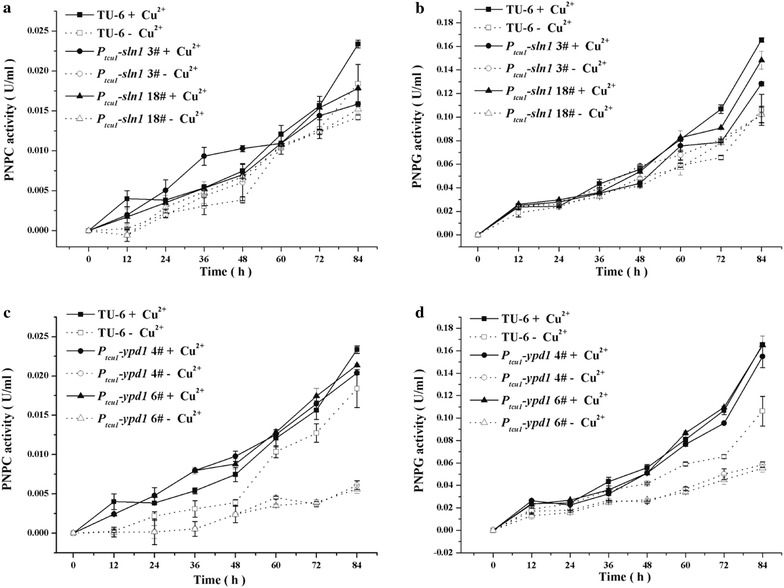
Here we established the physiological role of two upstream regulatory branches, the Sho1 branch and the Sln1 branch, of the Hog1-type Tmk3 pathway in . Deletion of of the Sho1 branch or repression of of the Sln1 branch reduced the resistance to high salt stress, whereas TrSho1 showed an opposing effect to that of TrSte20 and the identified TrSln1 seemed to be dispensable in the osmotic regulation. The Sho1 and Sln1 branches also participated in the cell wall integrity maintenance and other stress responses (i.e. oxidative and thermo stresses). Notably, TrSho1 and TrSte20 of the Sho1 branch and TrYpd1 of the Sln1 branch were shown to be differentially involved in the cellulase production of . Repression of hardly affected cellulase induction, whereas overexpression of resulted in the reduced production of cellulases. Contrary to the case of , repression of or deletion of significantly reduced the transcription of cellulase genes.
TrSho1 and TrSte20 of the Sho1 branch and TrYpd1 of the Sln1 branch are all involved in general stress responses including hyperosmotic regulation and cell wall integrity maintenance. Moreover, our study revealed that the Sho1 and Sln1 osmosensing pathways are differentially involved in the regulation of cellulase production in . The Sho1 branch positively regulated the production of cellulases and the transcription of cellulase genes while TrYpd1 of the Sln1 branch negatively controlled the cellulase production, supporting the crosstalks of osmosensing and nutrient sensing.
具有较高的蛋白质分泌能力,是工业中最重要的纤维素酶产生菌。然而,纤维素诱导过程中的外部信号感知和细胞内信号转导仍不清楚。作为所有真核生物中最普遍的信号转导途径之一,丝裂原活化蛋白激酶(MAPK)途径及其上游感知和信号成分参与包括应激和营养感知在内的各种生理过程。特别是,据报道Hog1型MAPK Tmk3参与了纤维素酶的产生。
在这里,我们确定了Hog1型Tmk3途径的两个上游调节分支,即Sho1分支和Sln1分支,在中的生理作用。Sho1分支的缺失或Sln1分支的抑制降低了对高盐胁迫的抗性,而TrSho1表现出与TrSte20相反的作用,并且所鉴定的TrSln1在渗透调节中似乎是可有可无的。Sho1和Sln1分支也参与了细胞壁完整性维持和其他应激反应(即氧化应激和热应激)。值得注意的是,Sho1分支的TrSho1和TrSte20以及Sln1分支的TrYpd1被证明在纤维素酶的产生中发挥不同作用。的抑制几乎不影响纤维素酶诱导,而的过表达导致纤维素酶产量降低。与情况相反,的抑制或的缺失显著降低了纤维素酶基因的转录。
Sho1分支的TrSho1和TrSte20以及Sln1分支的TrYpd1都参与了包括高渗调节和细胞壁完整性维持在内的一般应激反应。此外,我们的研究表明,Sho1和Sln1渗透感应途径在纤维素酶产生的调节中发挥不同作用。Sho1分支正向调节纤维素酶的产生和纤维素酶基因的转录,而Sln1分支的TrYpd1负向控制纤维素酶的产生,支持渗透感应和营养感应的相互作用。