Department of Biochemistry and Molecular Genetics, College of Medicine, University of Illinois at Chicago, Chicago, United States.
Research & Development Section, Jesse Brown VA Medical Center, Chicago, United States.
Elife. 2018 Apr 24;7:e32213. doi: 10.7554/eLife.32213.
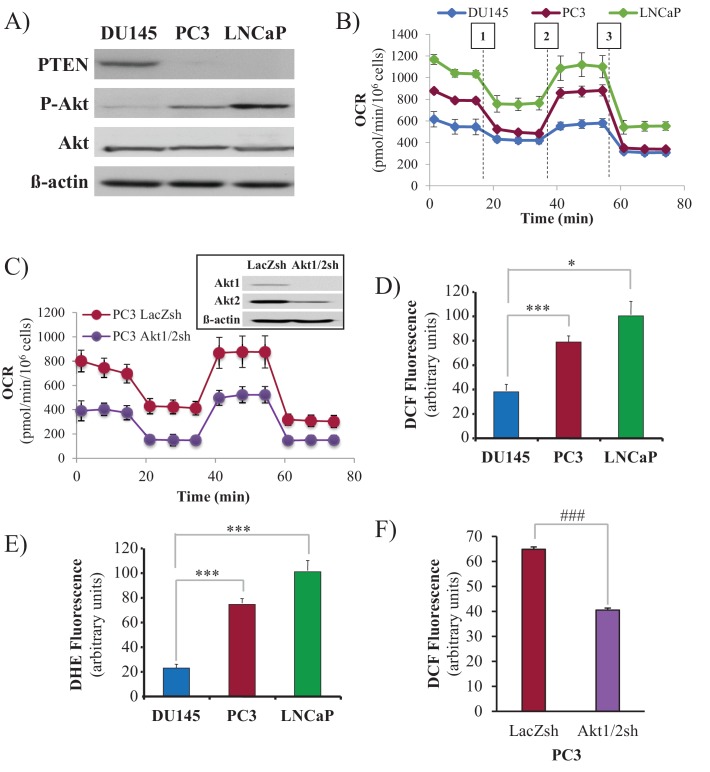
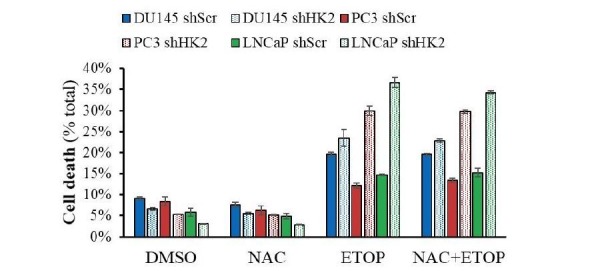
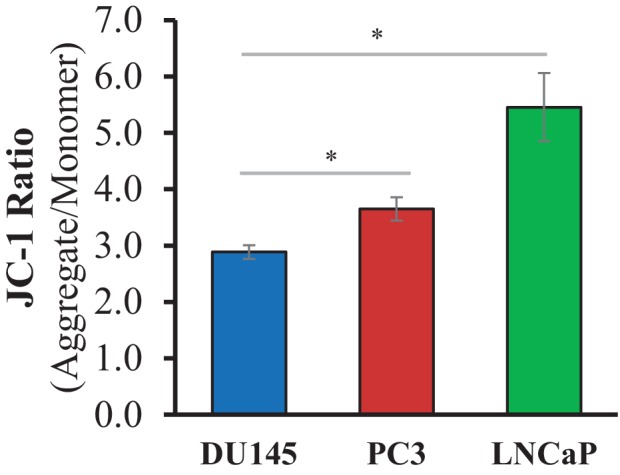
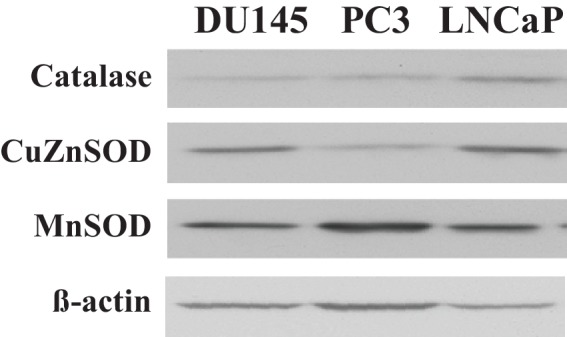
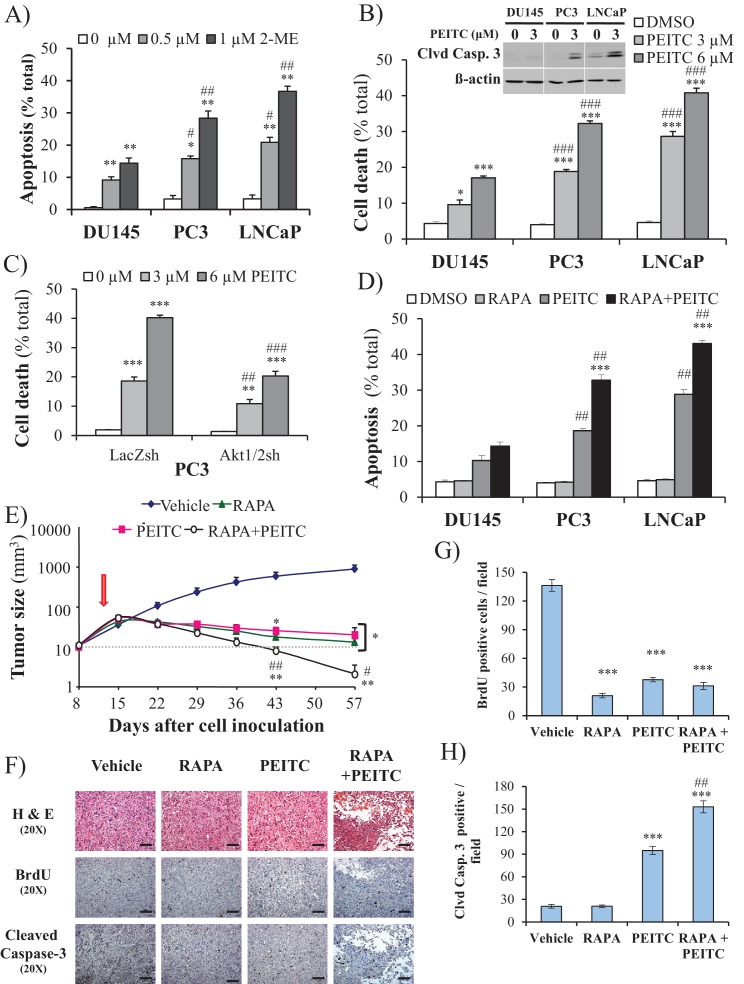
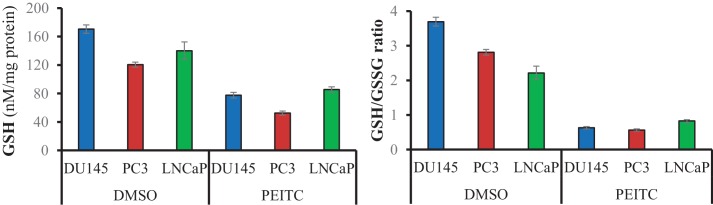
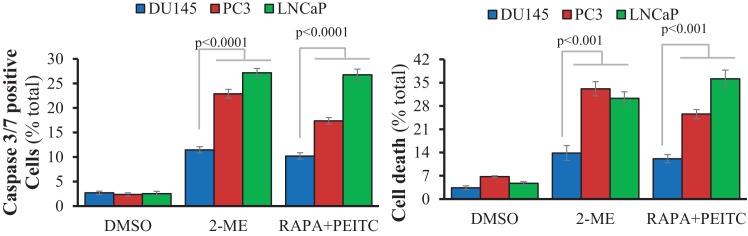
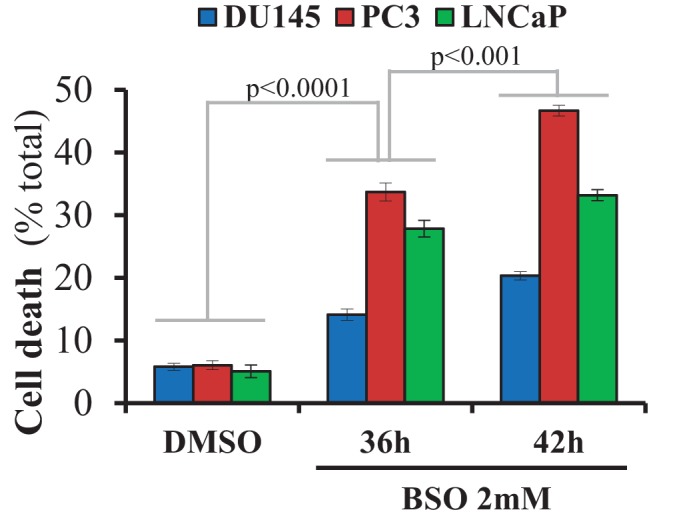
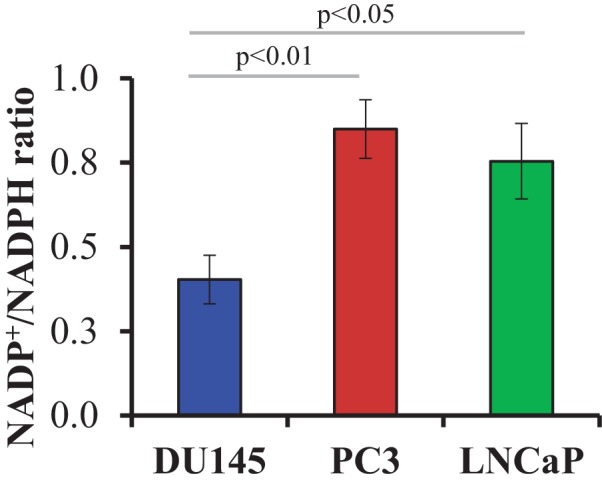
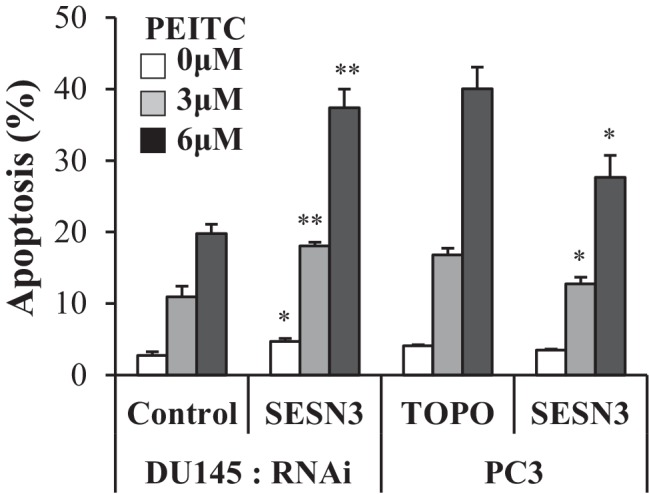
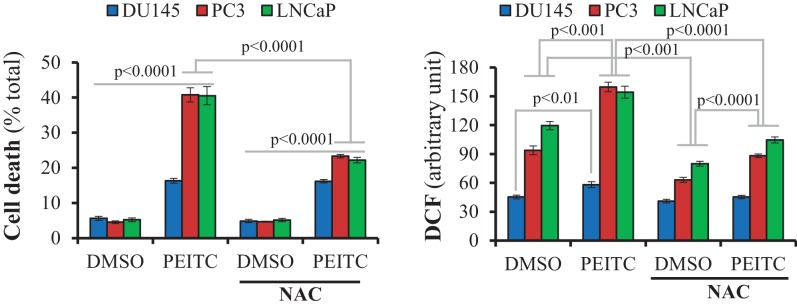
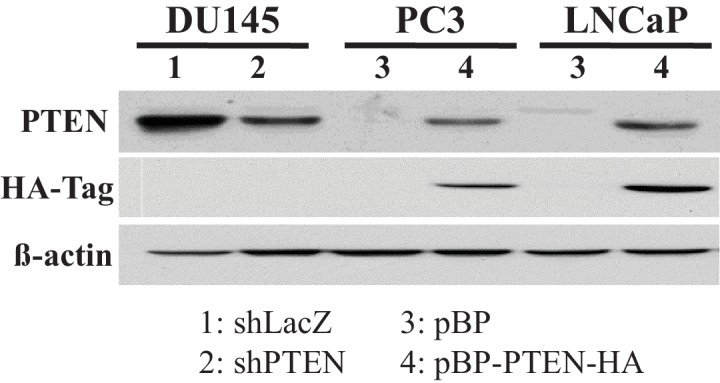
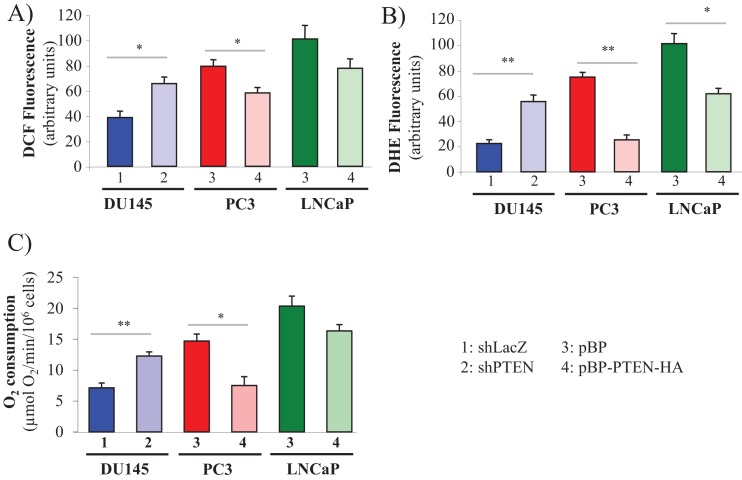
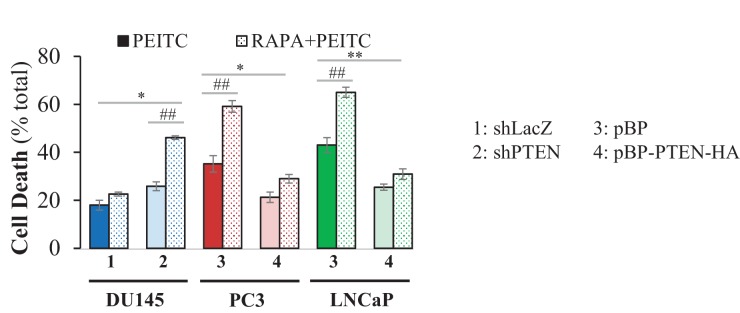
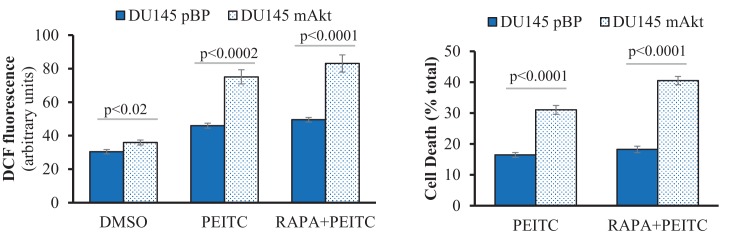
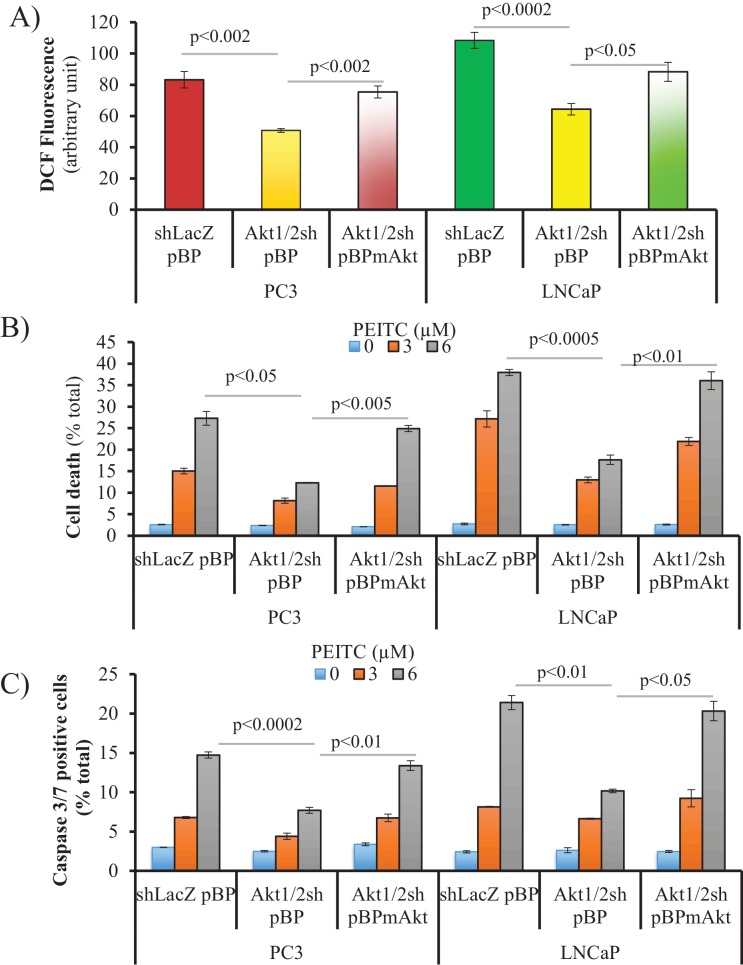
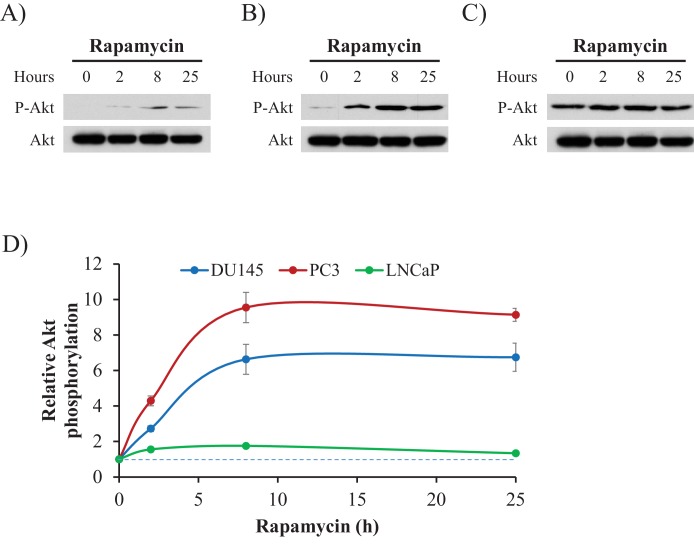
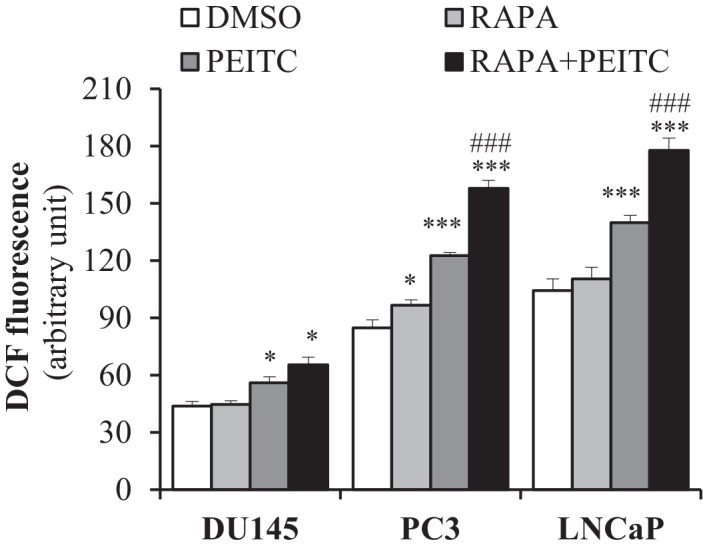
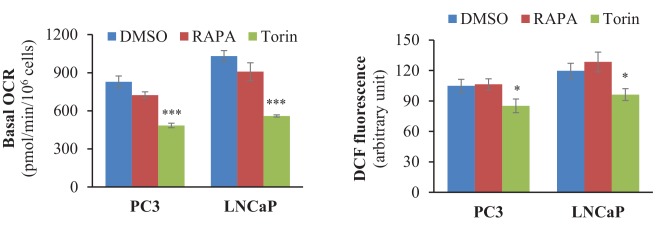
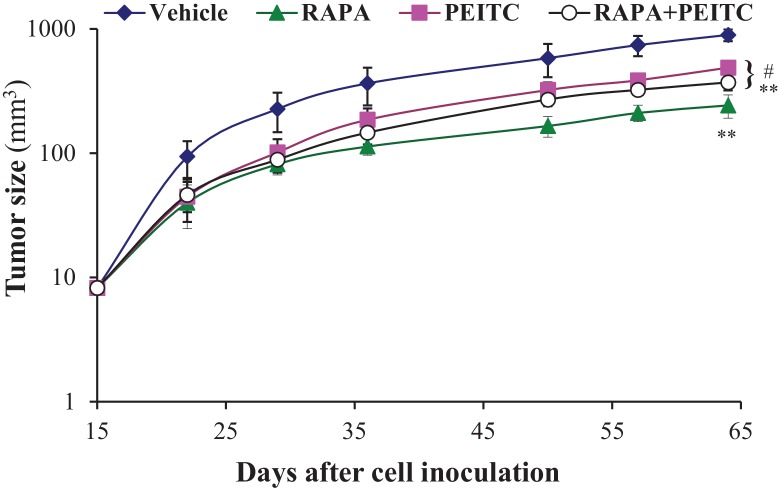
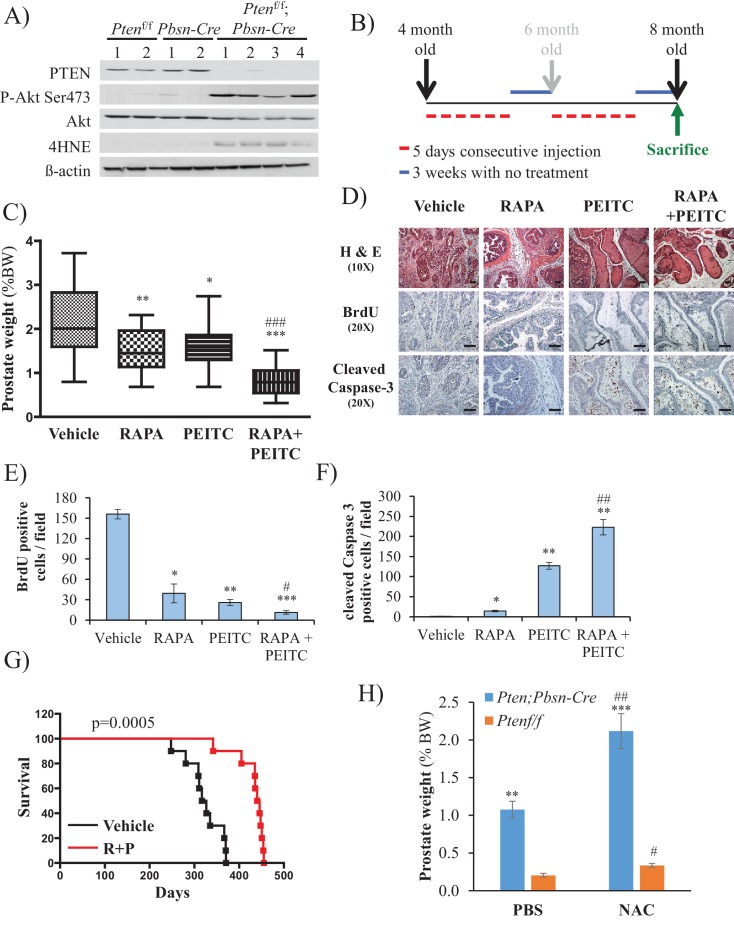
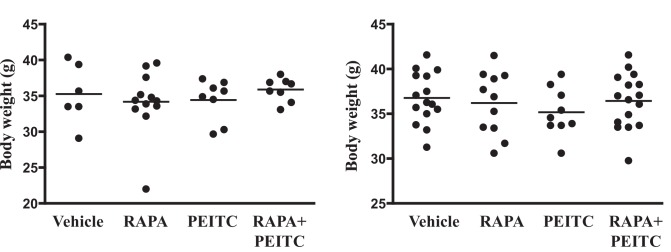
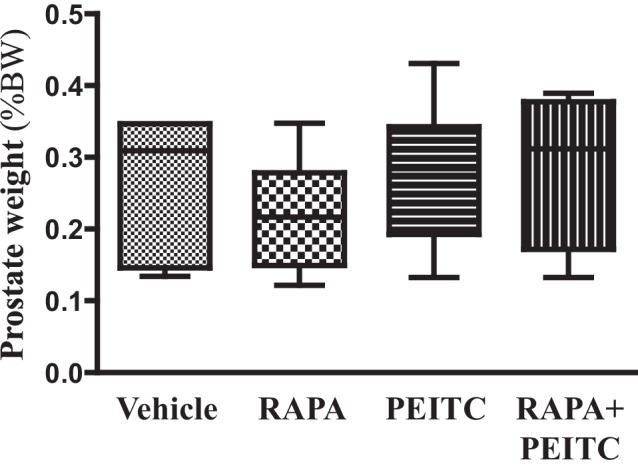
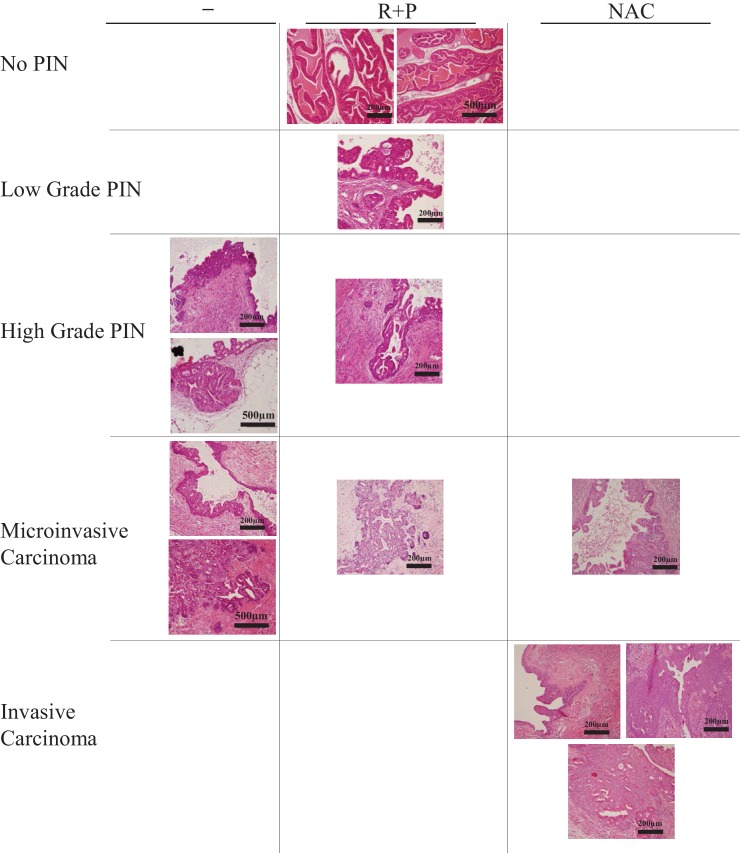
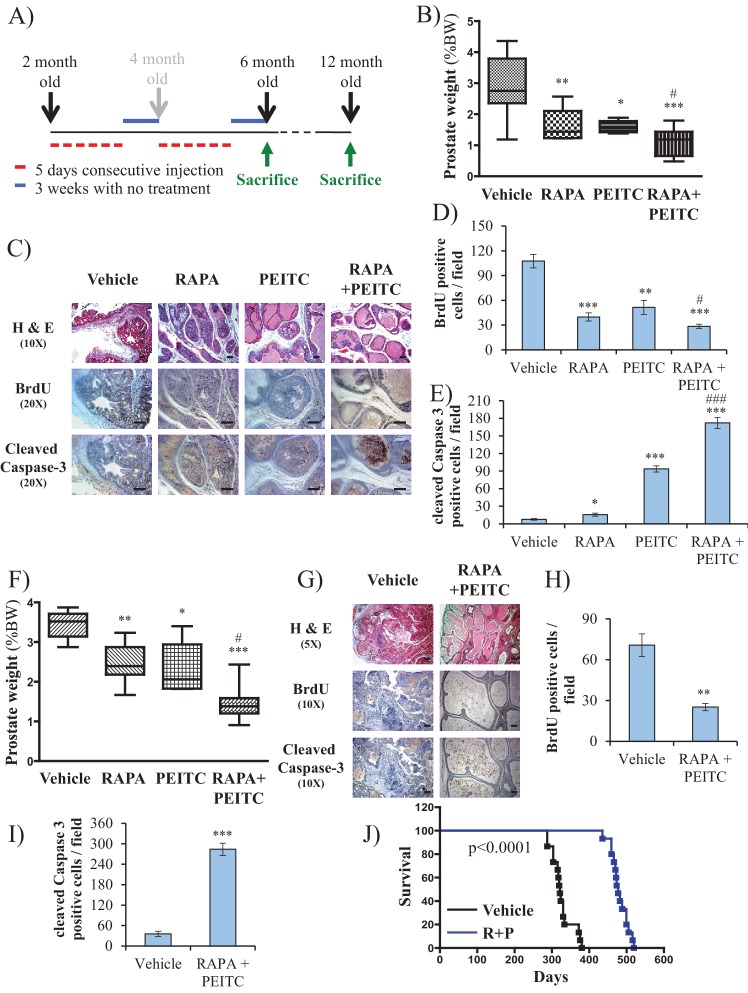
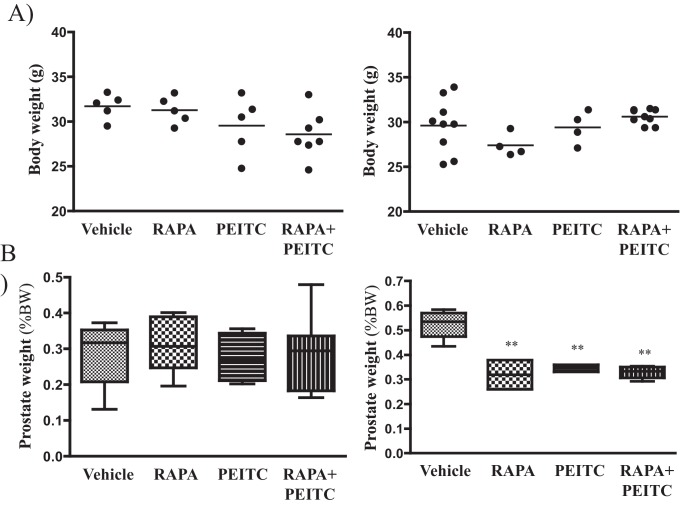
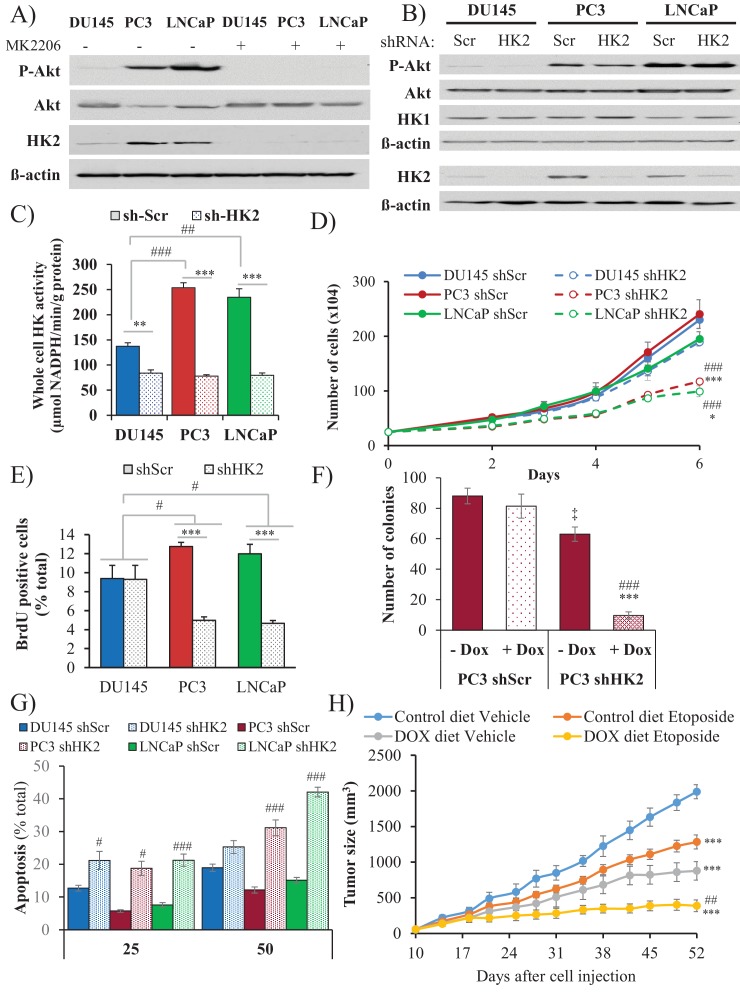
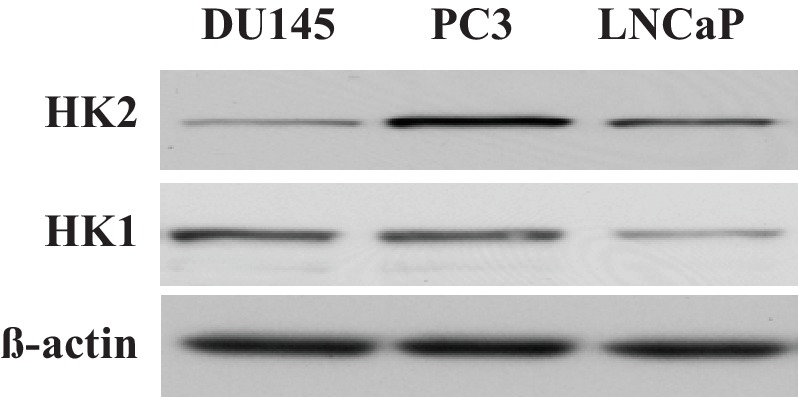
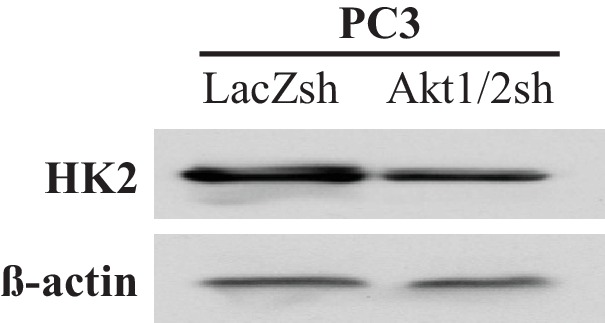
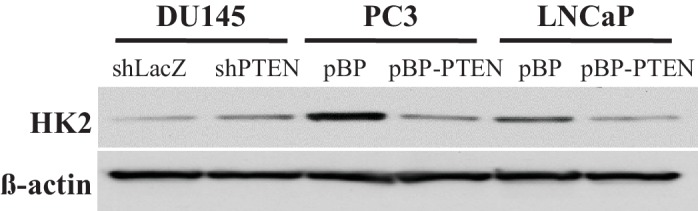
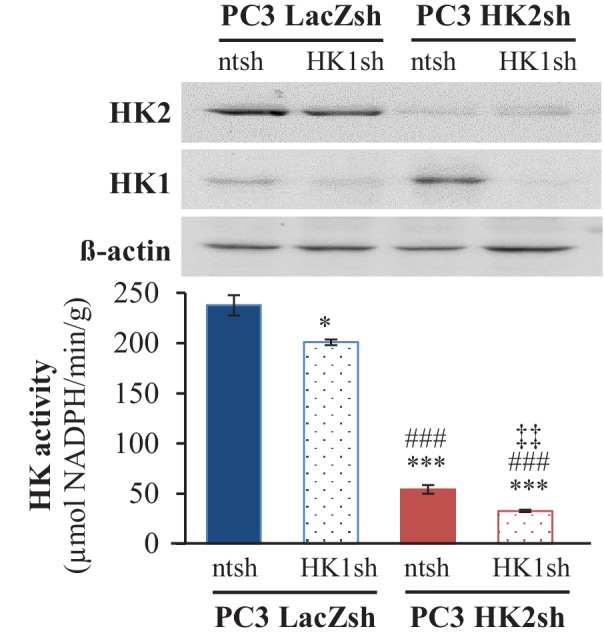
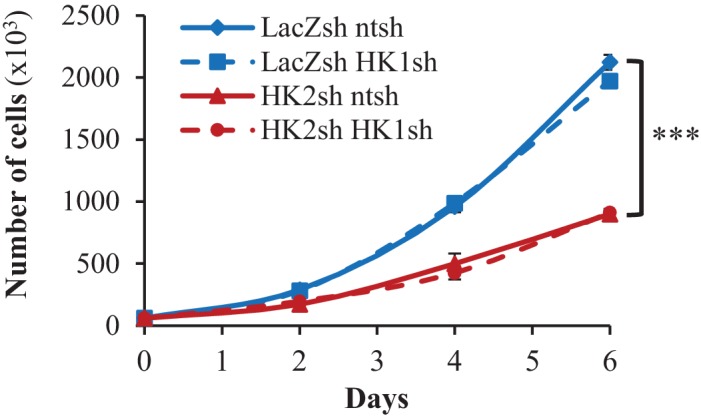
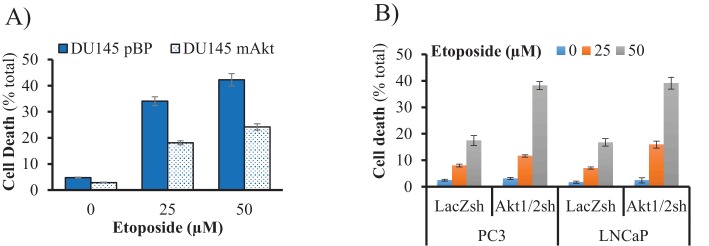
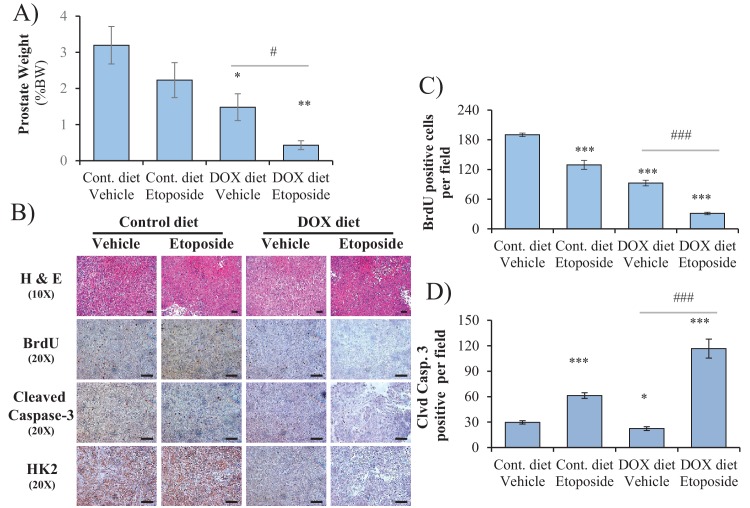
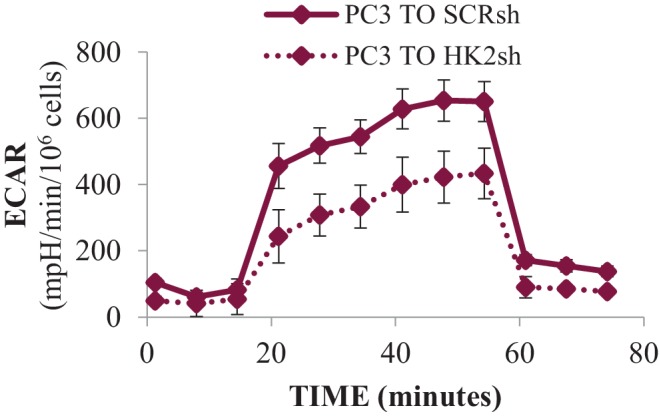
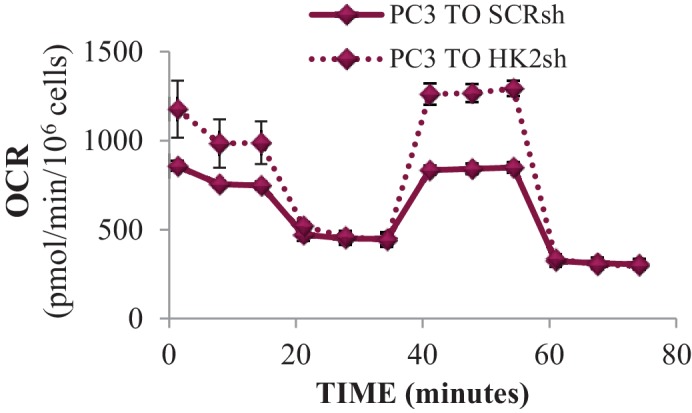
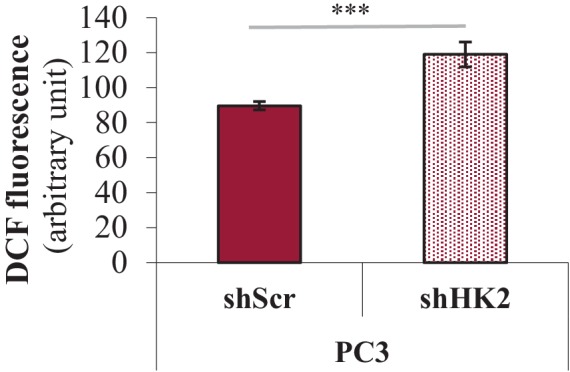
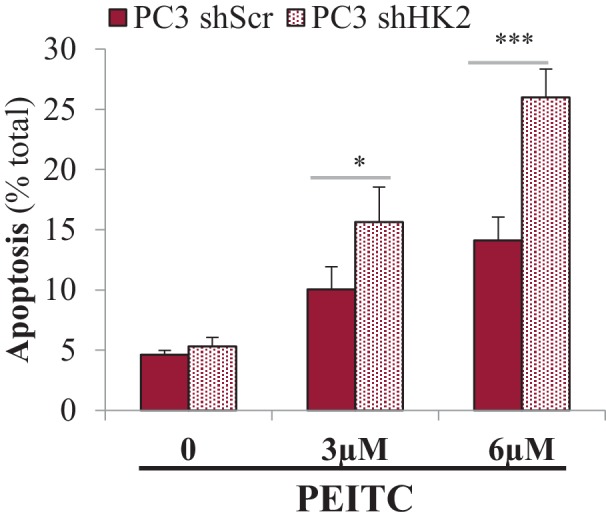
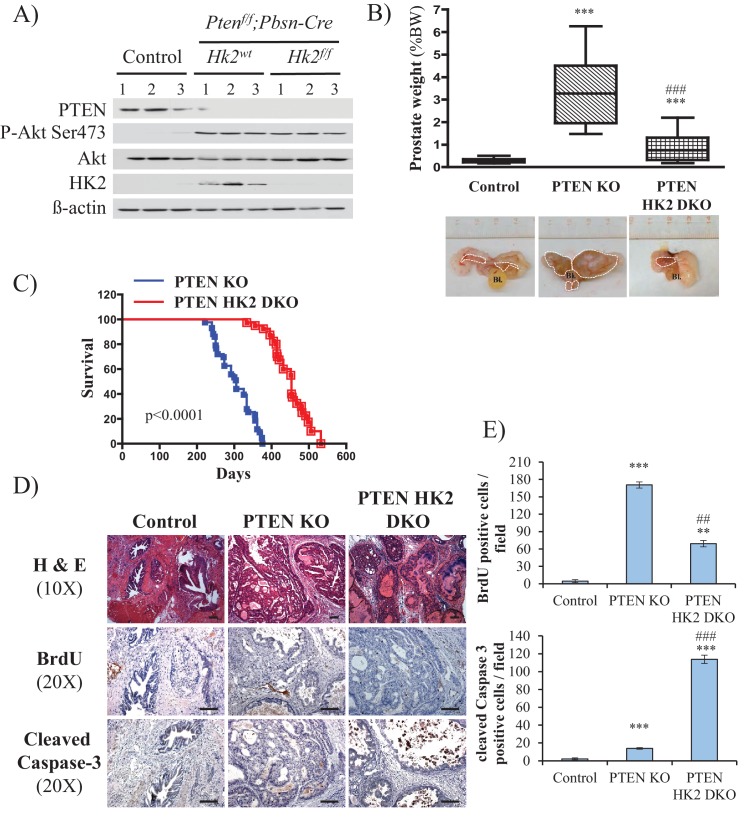
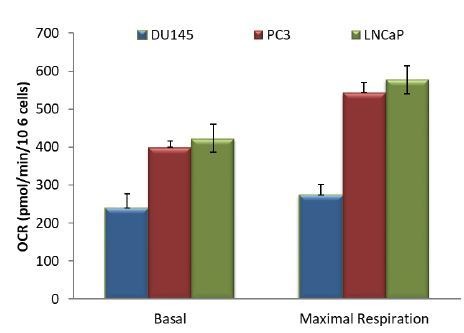
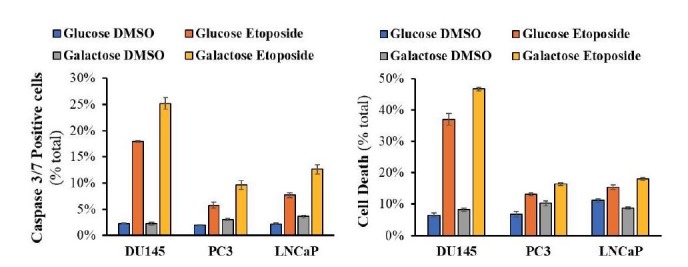
Akt activation in human cancers exerts chemoresistance, but pan-Akt inhibition elicits adverse consequences. We exploited the consequences of Akt-mediated mitochondrial and glucose metabolism to selectively eradicate and evade chemoresistance of prostate cancer displaying hyperactive Akt. PTEN-deficient prostate cancer cells that display hyperactivated Akt have high intracellular reactive oxygen species (ROS) levels, in part, because of Akt-dependent increase of oxidative phosphorylation. High intracellular ROS levels selectively sensitize cells displaying hyperactive Akt to ROS-induced cell death enabling a therapeutic strategy combining a ROS inducer and rapamycin in PTEN-deficient prostate tumors in mouse models. This strategy elicited tumor regression, and markedly increased survival even after the treatment was stopped. By contrast, exposure to antioxidant increased prostate tumor progression. To increase glucose metabolism, Akt activation phosphorylated HK2 and induced its expression. Indeed, HK2 deficiency in mouse models of Pten-deficient prostate cancer elicited a marked inhibition of tumor development and extended lifespan.
在人类癌症中,Akt 的激活会产生化疗耐药性,但泛 Akt 抑制会引发不良反应。我们利用 Akt 介导的线粒体和葡萄糖代谢的后果,选择性地根除和逃避 Akt 过度激活的前列腺癌的化疗耐药性。PTEN 缺陷型前列腺癌细胞中 Akt 过度激活会导致细胞内活性氧(ROS)水平升高,部分原因是 Akt 依赖性氧化磷酸化增加。高细胞内 ROS 水平选择性地使 Akt 过度激活的细胞对 ROS 诱导的细胞死亡敏感,从而使在小鼠模型中的 PTEN 缺陷型前列腺肿瘤中联合使用 ROS 诱导剂和雷帕霉素成为一种治疗策略。这种策略引发了肿瘤消退,并显著提高了即使在治疗停止后的存活率。相比之下,暴露于抗氧化剂会增加前列腺肿瘤的进展。为了增加葡萄糖代谢,Akt 的激活磷酸化 HK2 并诱导其表达。事实上,在 Pten 缺陷型前列腺癌的小鼠模型中,HK2 缺乏会显著抑制肿瘤的发展并延长寿命。