Wellcome Centre for Cell Biology, School of Biological Sciences, University of Edinburgh, Edinburgh, United Kingdom.
Advanced ICT Research Institute Kobe, National Institute of Information and Communications Technology, Kobe, Japan.
Elife. 2018 May 29;7:e33465. doi: 10.7554/eLife.33465.
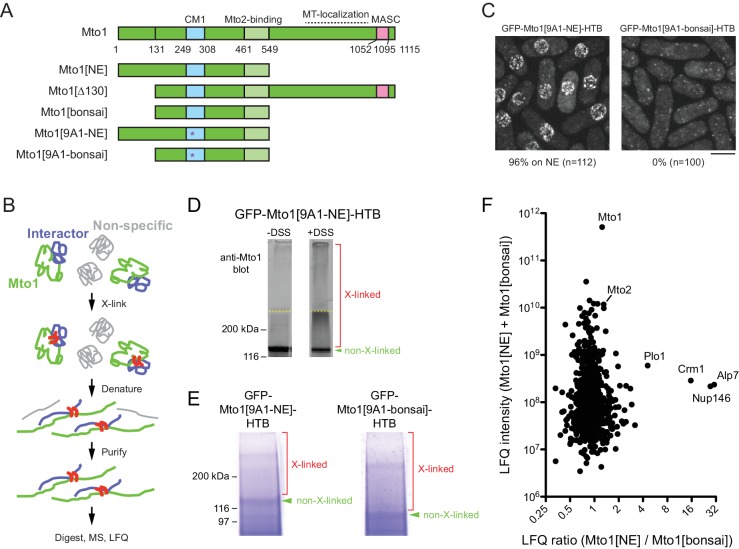
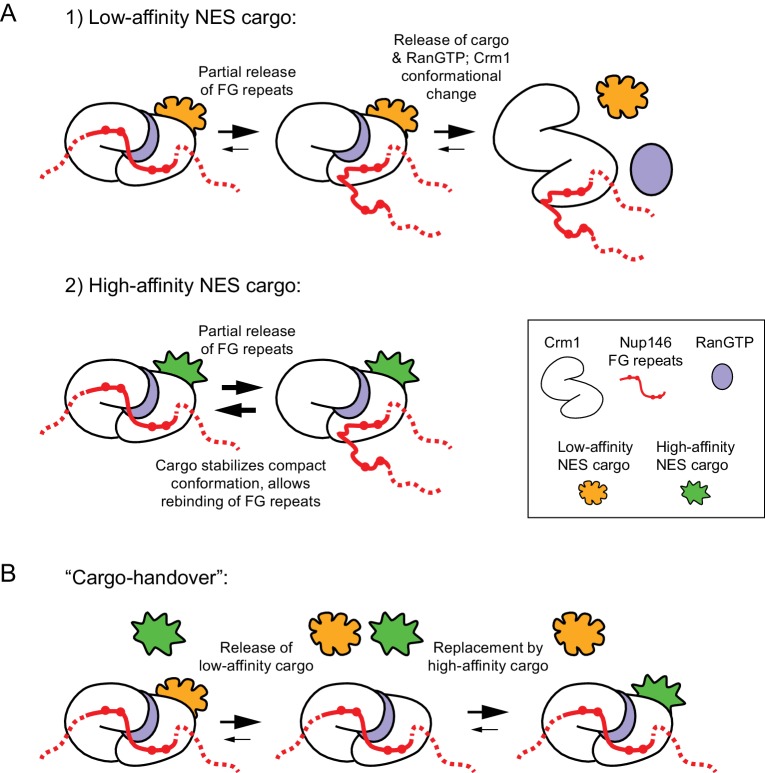
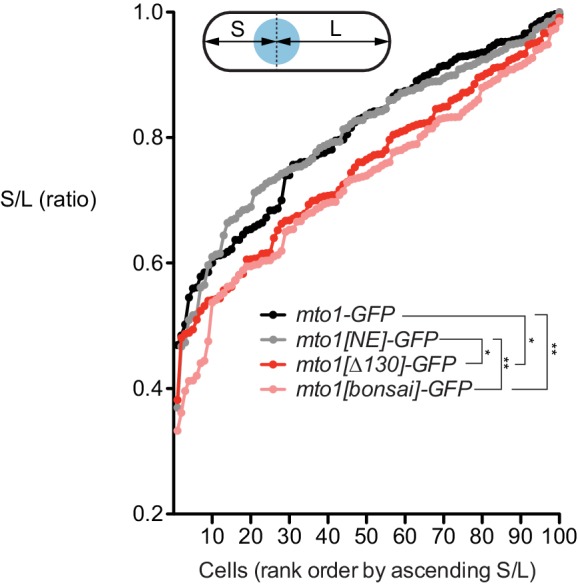
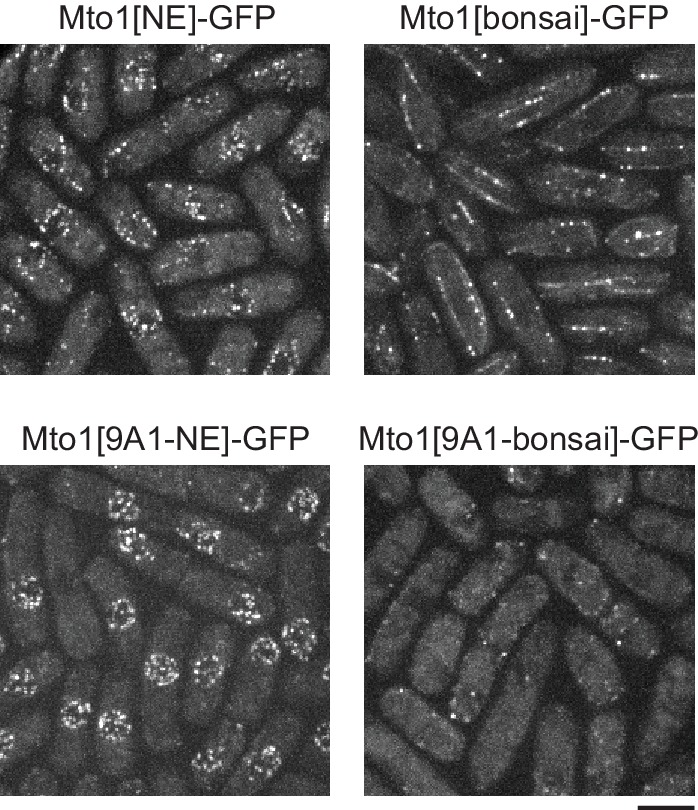
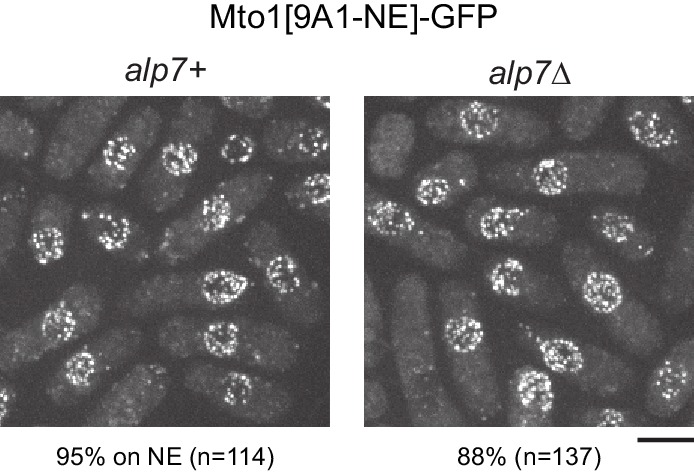
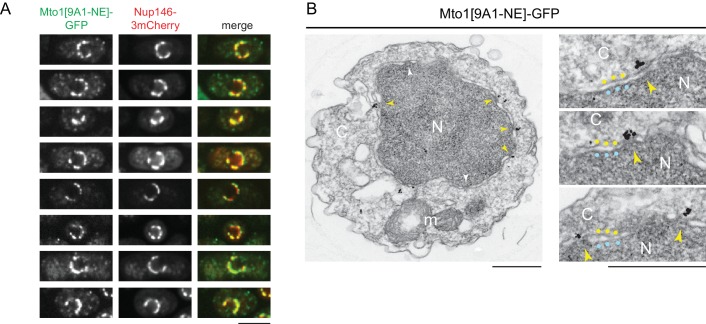
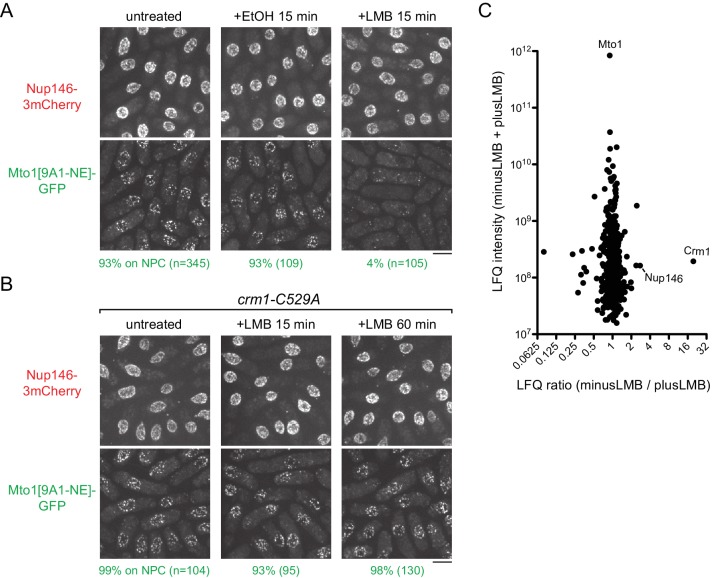
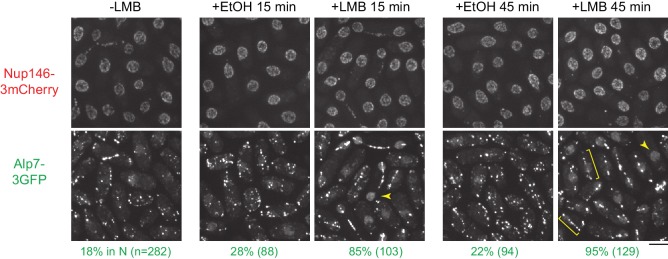
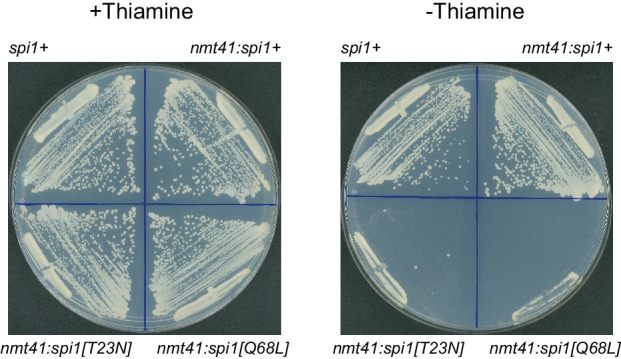
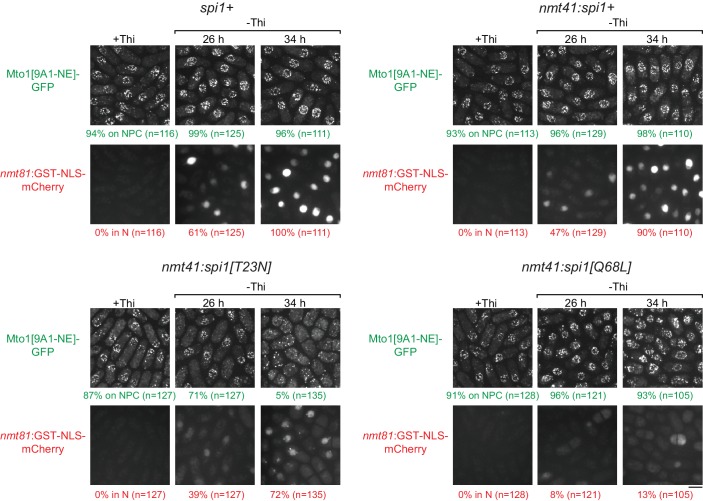
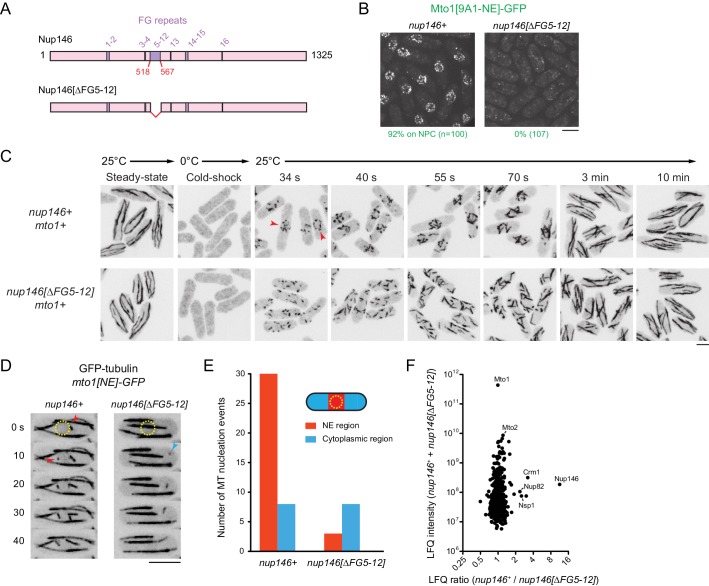
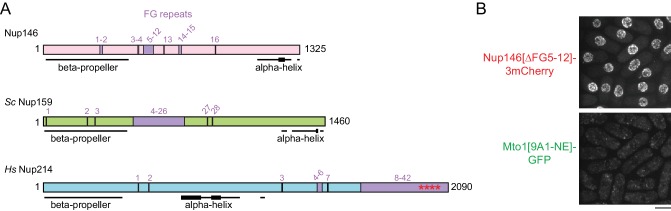
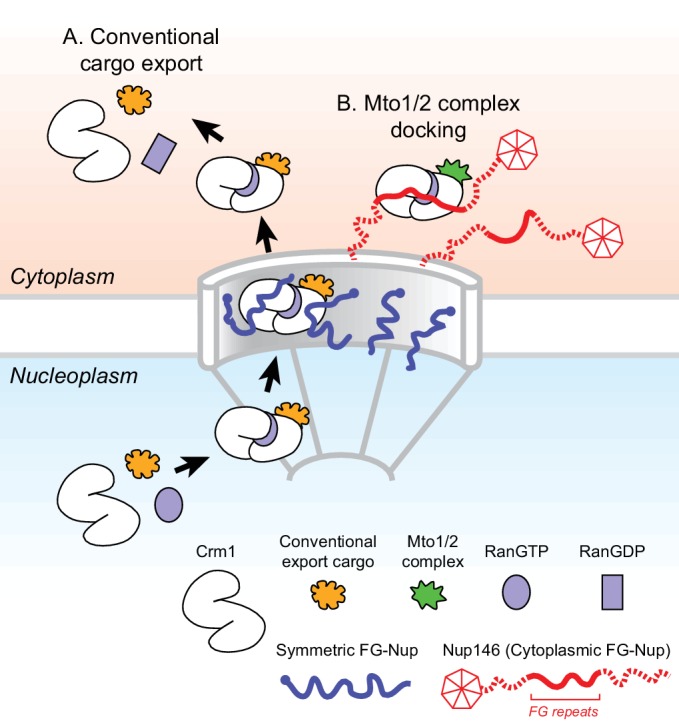
Non-centrosomal microtubule organizing centers (MTOCs) are important for microtubule organization in many cell types. In fission yeast , the protein Mto1, together with partner protein Mto2 (Mto1/2 complex), recruits the γ-tubulin complex to multiple non-centrosomal MTOCs, including the nuclear envelope (NE). Here, we develop a comparative-interactome mass spectrometry approach to determine how Mto1 localizes to the NE. Surprisingly, we find that Mto1, a constitutively cytoplasmic protein, docks at nuclear pore complexes (NPCs), via interaction with exportin Crm1 and cytoplasmic FG-nucleoporin Nup146. Although Mto1 is not a nuclear export cargo, it binds Crm1 via a nuclear export signal-like sequence, and docking requires both Ran in the GTP-bound state and Nup146 FG repeats. In addition to determining the mechanism of MTOC formation at the NE, our results reveal a novel role for Crm1 and the nuclear export machinery in the stable docking of a cytoplasmic protein complex at NPCs.
非中心体微管组织中心(MTOC)对于许多细胞类型的微管组织都很重要。在裂殖酵母中,蛋白质 Mto1 与伴侣蛋白 Mto2(Mto1/2 复合物)一起将 γ-微管蛋白复合物招募到多个非中心体 MTOC,包括核膜(NE)。在这里,我们开发了一种比较相互作用组质谱法来确定 Mto1 如何定位到 NE。令人惊讶的是,我们发现 Mto1,一种组成型细胞质蛋白,通过与输出蛋白 Crm1 和细胞质 FG-核孔蛋白 Nup146 的相互作用,停靠在核孔复合物(NPC)上。尽管 Mto1 不是核输出货物,但它通过类似于核输出信号的序列与 Crm1 结合,并且对接需要 GTP 结合状态下的 Ran 和 Nup146 FG 重复序列。除了确定 NE 处 MTOC 形成的机制外,我们的结果还揭示了 Crm1 和核输出机制在细胞质蛋白复合物在 NPC 上的稳定停靠中的新作用。