Department of Molecular and Cellular Physiology, Stanford University, Stanford, United States.
Department of Neurology and Neurological Sciences, Stanford University, Stanford, United States.
Elife. 2018 Jul 9;7:e36497. doi: 10.7554/eLife.36497.
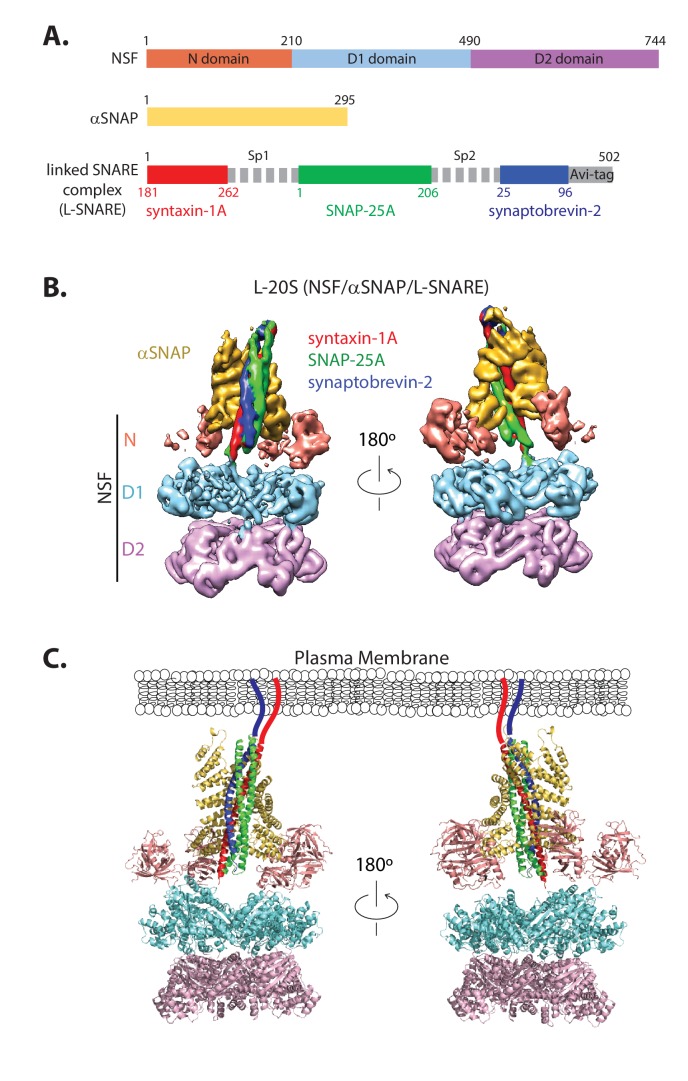
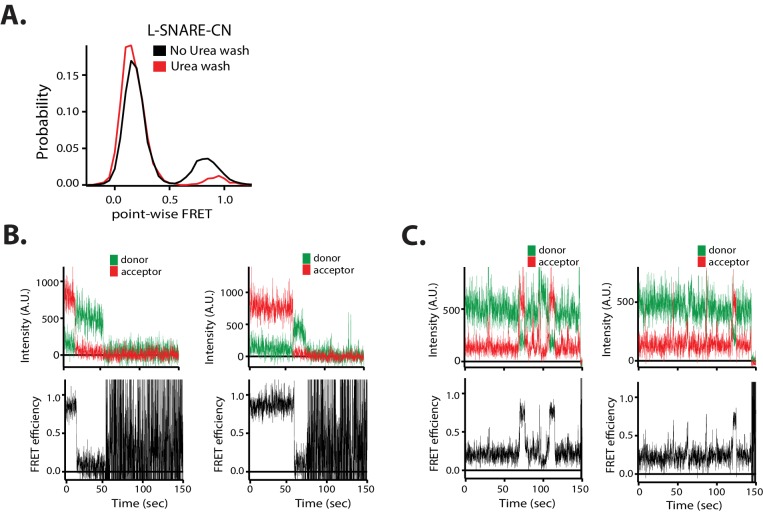
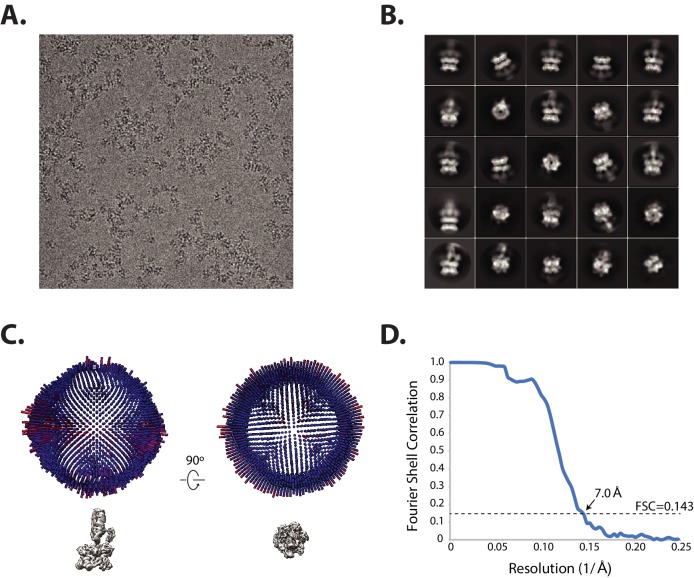
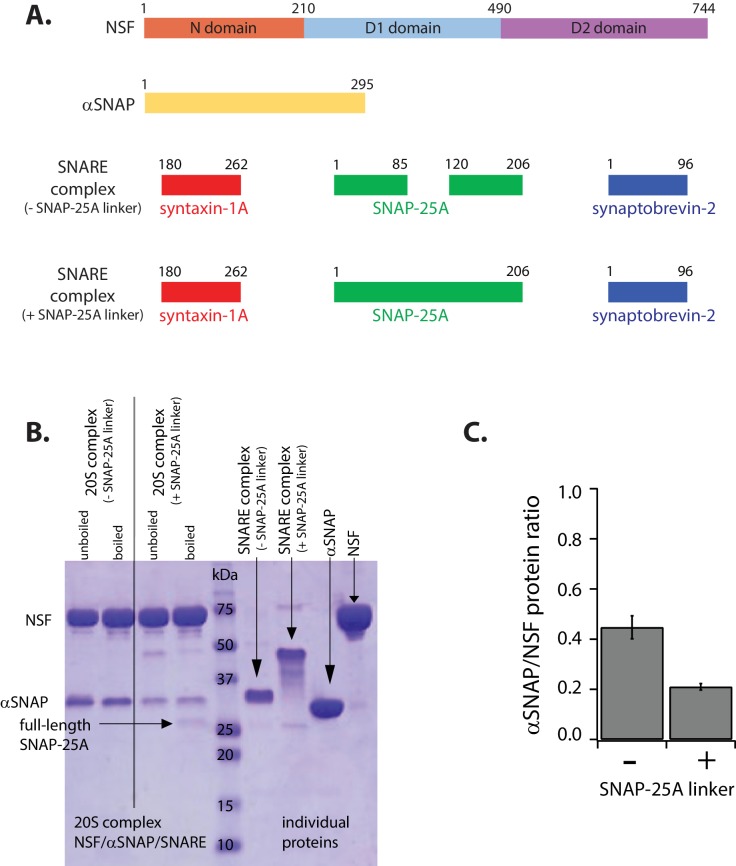
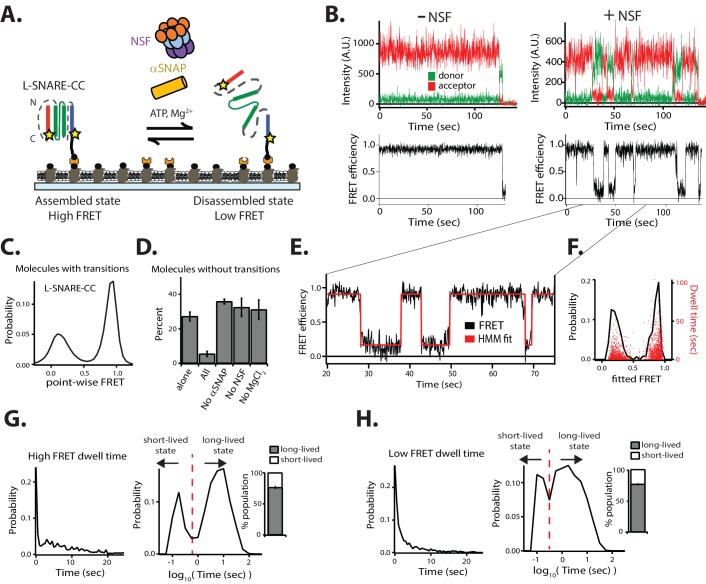
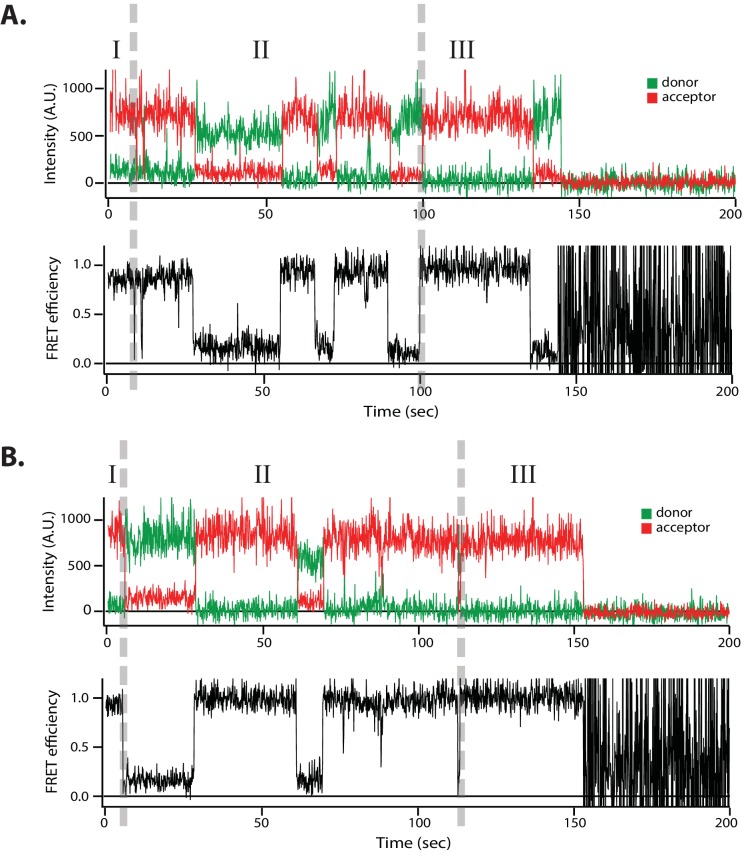
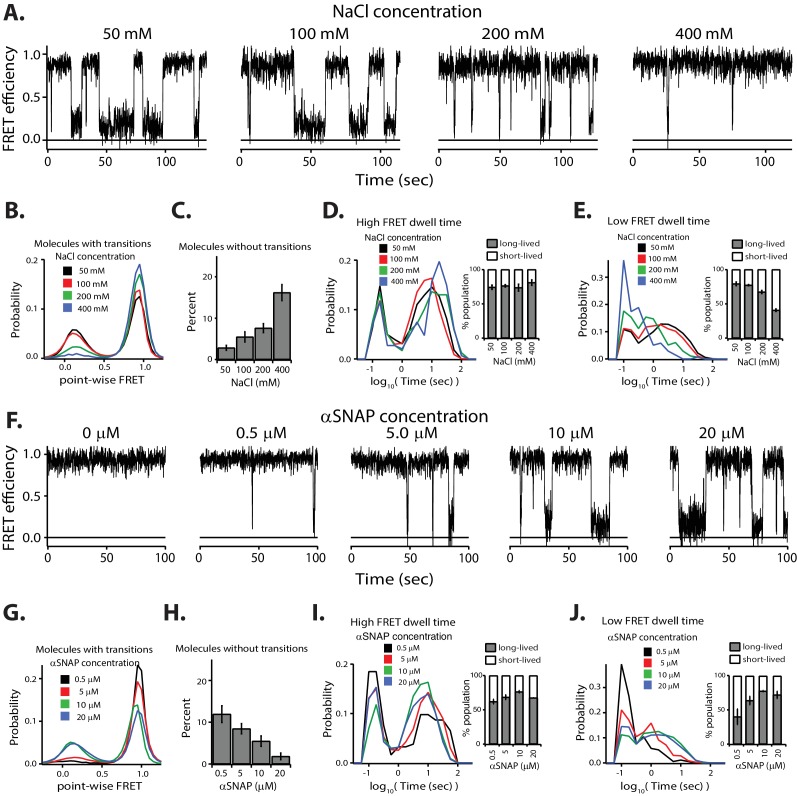
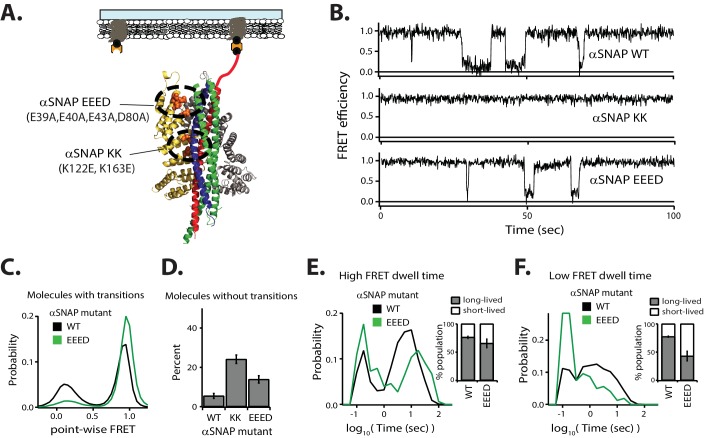
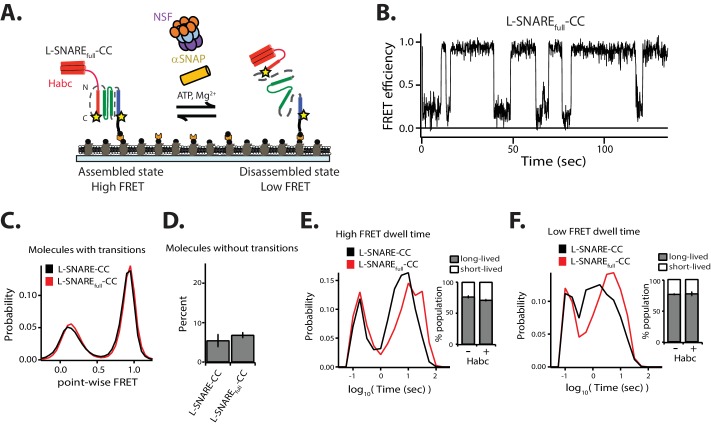
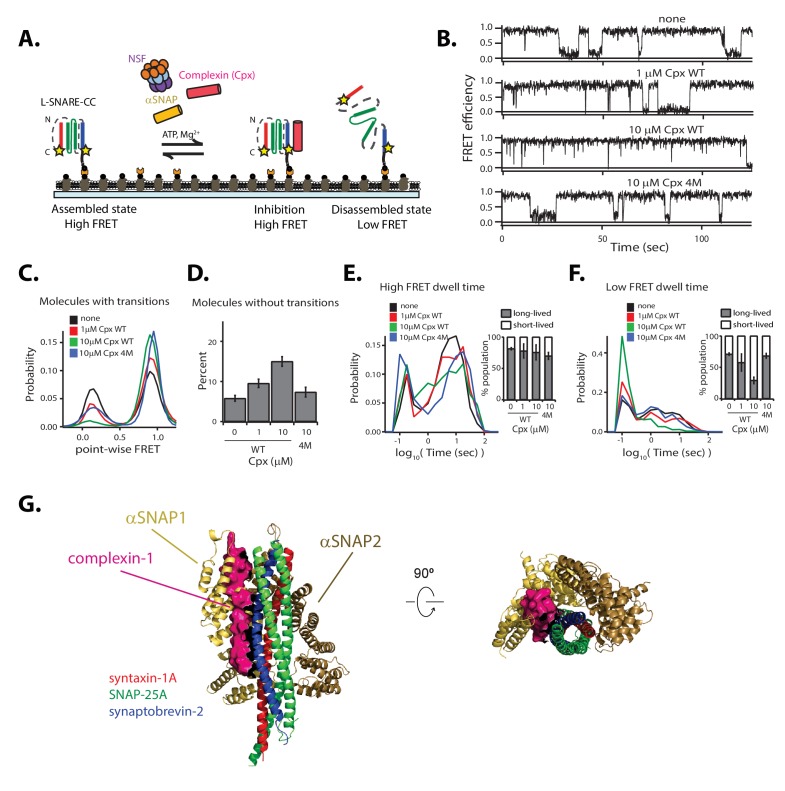
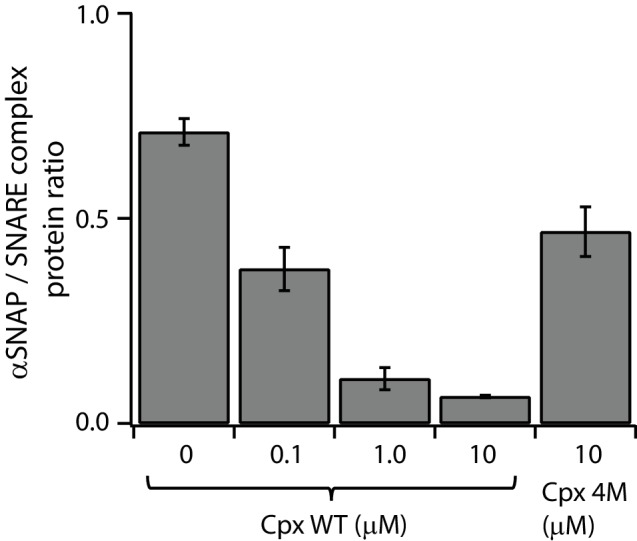
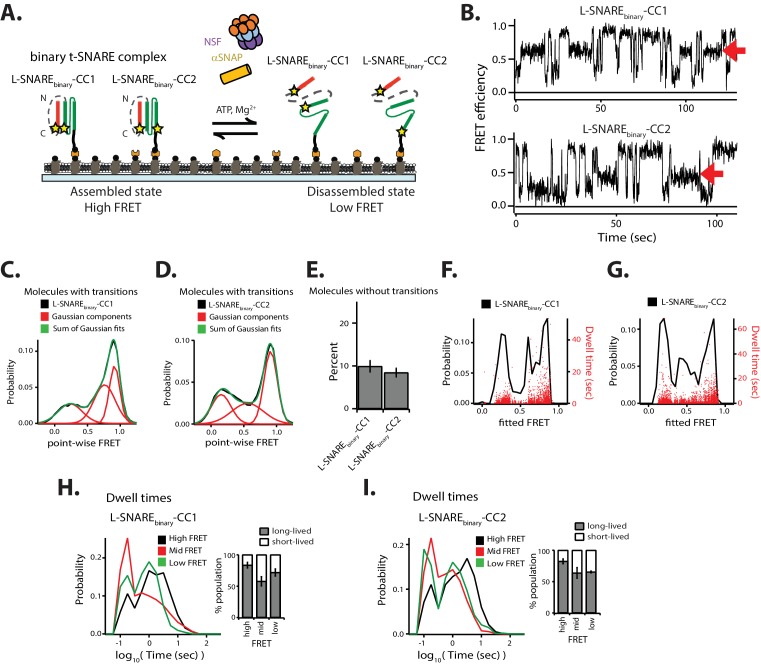
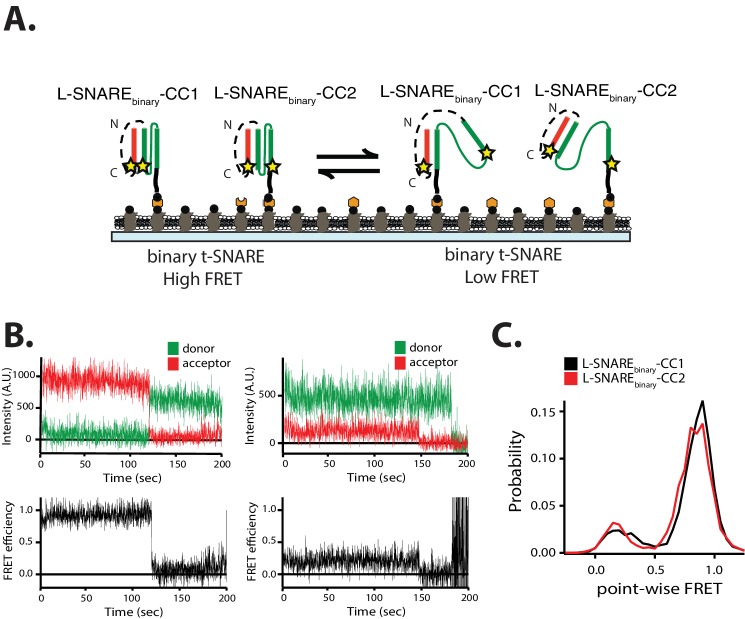
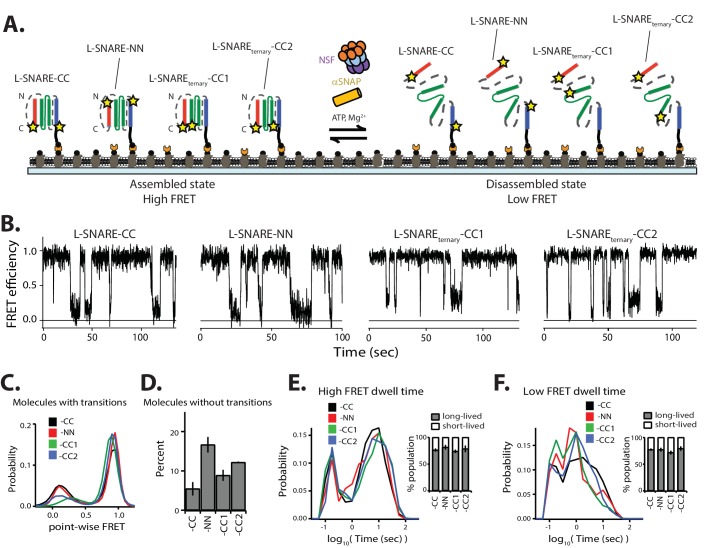
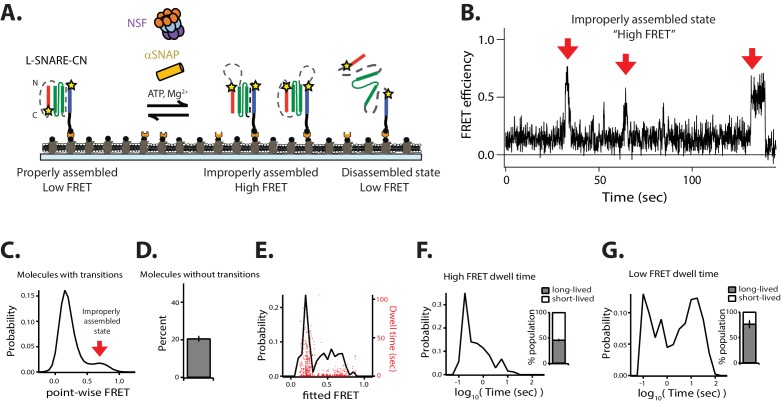
SNARE complex disassembly by the ATPase NSF is essential for neurotransmitter release and other membrane trafficking processes. We developed a single-molecule FRET assay to monitor repeated rounds of NSF-mediated disassembly and reassembly of individual SNARE complexes. For ternary neuronal SNARE complexes, disassembly proceeds in a single step within 100 msec. We observed short- (<0.32 s) and long-lived (≥0.32 s) disassembled states. The long-lived states represent fully disassembled SNARE complex, while the short-lived states correspond to failed disassembly or immediate reassembly. Either high ionic strength or decreased αSNAP concentration reduces the disassembly rate while increasing the frequency of short-lived states. NSF is also capable of disassembling anti-parallel ternary SNARE complexes, implicating it in quality control. Finally, complexin-1 competes with αSNAP binding to the SNARE complex; addition of complexin-1 has an effect similar to that of decreasing the αSNAP concentration, possibly differentially regulating cis and trans SNARE complexes disassembly.
SNARE 复合物的 ATP 酶 NSF 解体对于神经递质释放和其他膜运输过程至关重要。我们开发了一种单分子 FRET 测定法来监测 NSF 介导的单个 SNARE 复合物的重复解体和组装。对于三元神经元 SNARE 复合物,在 100 毫秒内可进行单步解体。我们观察到短(<0.32 秒)和长寿命(≥0.32 秒)的解体状态。长寿命状态代表完全解体的 SNARE 复合物,而短寿命状态则对应于失败的解体或立即重新组装。高离子强度或降低 αSNAP 浓度会降低解体速度,同时增加短寿命状态的频率。NSF 还能够解体反平行的三元 SNARE 复合物,暗示它参与了质量控制。最后,复合蛋白-1 与 αSNAP 竞争与 SNARE 复合物的结合;添加复合蛋白-1的效果类似于降低 αSNAP 浓度,可能会以不同的方式调节顺式和反式 SNARE 复合物的解体。