Perez Jimenez Tania E, Mealey Katrina L, Schnider Darren, Grubb Tamara L, Greene Stephen A, Court Michael H
Program in Individualized Medicine (PrIMe), Department of Veterinary Clinical Sciences, Pharmacogenomics Laboratory, Washington State University College of Veterinary Medicine, Pullman, Washington.
J Vet Pharmacol Ther. 2018 Dec;41(6):815-824. doi: 10.1111/jvp.12706. Epub 2018 Aug 16.
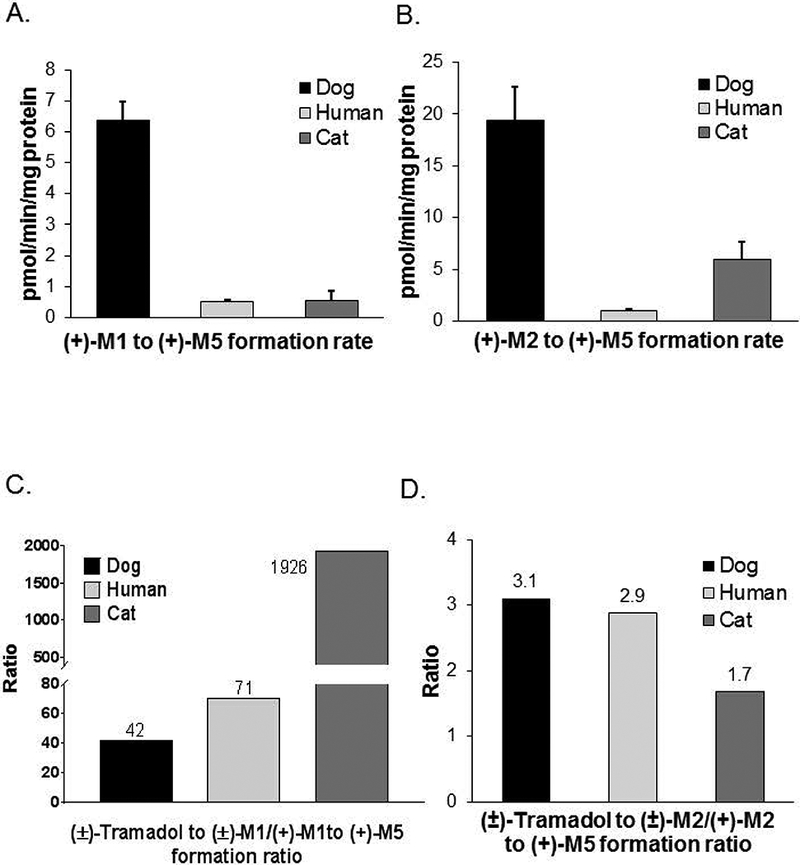
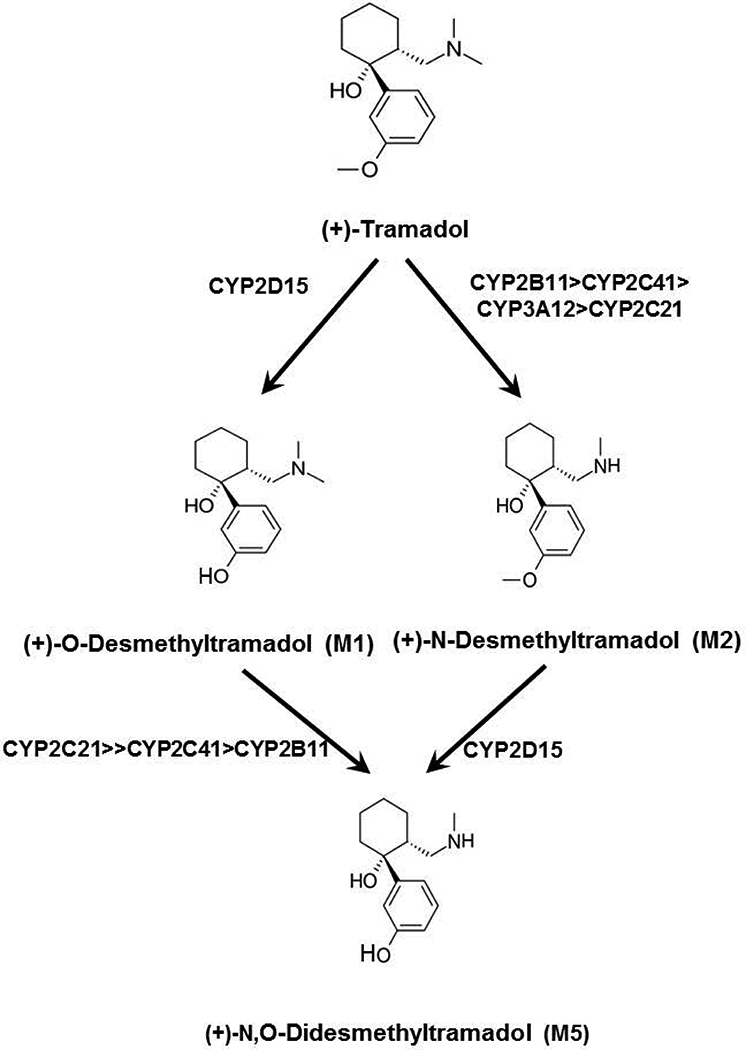
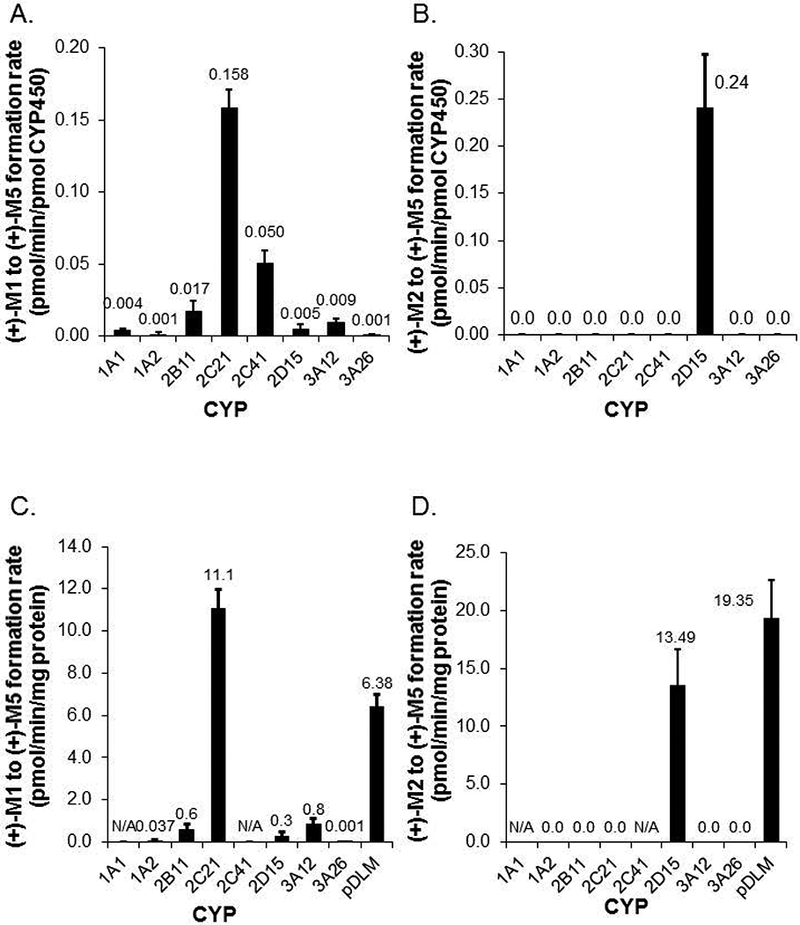
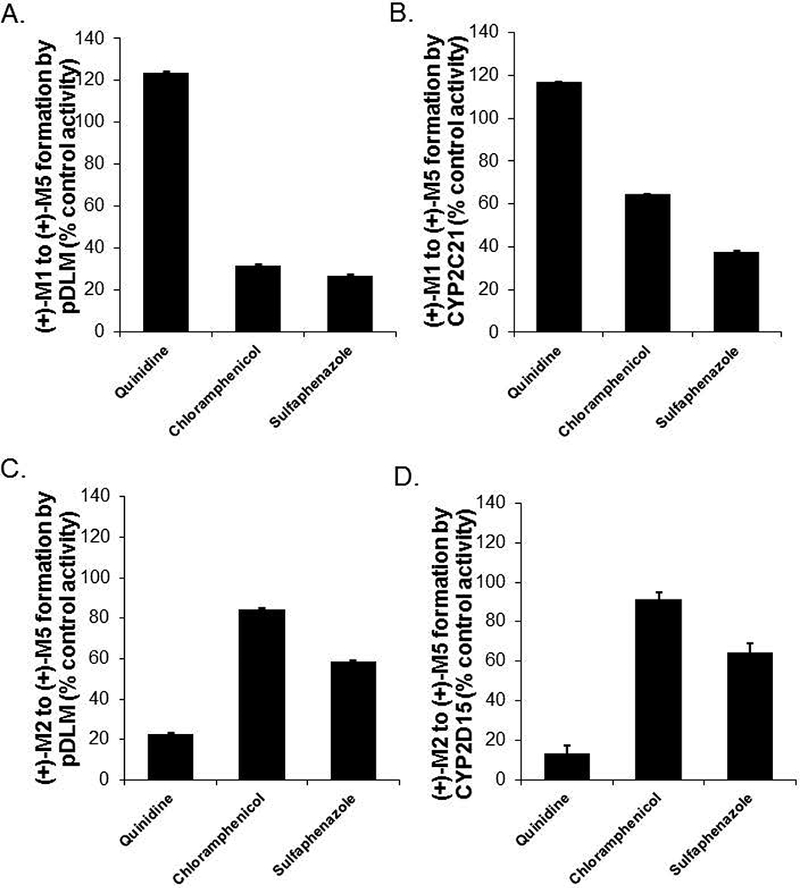
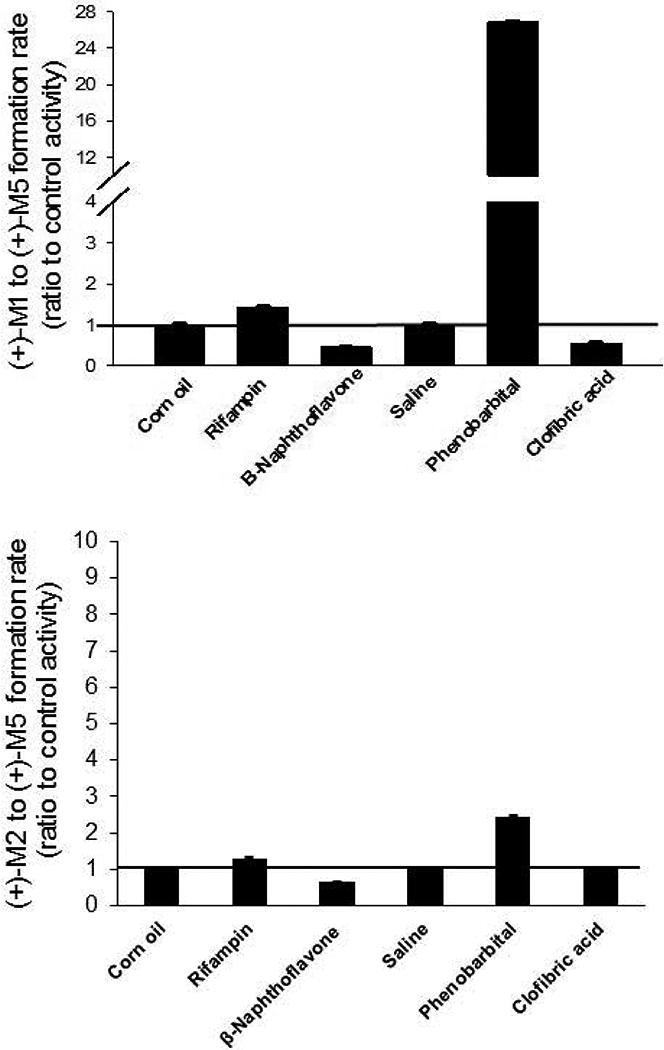
We previously showed that (+)-tramadol is metabolized in dog liver to (+)-M1 exclusively by CYP2D15 and to (+)-M2 by multiple CYPs, but primarily CYP2B11. However, (+)-M1 and (+)-M2 are further metabolized in dogs to (+)-M5, which is the major metabolite found in dog plasma and urine. In this study, we identified canine CYPs involved in metabolizing (+)-M1 and (+)-M2 using recombinant enzymes, untreated dog liver microsomes (DLMs), inhibitor-treated DLMs, and DLMs from CYP inducer-treated dogs. A canine P-glycoprotein expressing cell line was also used to evaluate whether (+)-tramadol, (+)-M1, (+)-M2, or (+)-M5 are substrates of canine P-glycoprotein, thereby limiting their distribution into the central nervous system. (+)-M5 was largely formed from (+)-M1 by recombinant CYP2C21 with minor contributions from CYP2C41 and CYP2B11. (+)-M5 formation in DLMs from (+)-M1 was potently inhibited by sulfaphenazole (CYP2C inhibitor) and chloramphenicol (CYP2B11 inhibitor) and was greatly increased in DLMs from phenobarbital-treated dogs. (+)-M5 was formed from (+)-M2 predominantly by CYP2D15. (+)-M5 formation from (+)-M1 in DLMs was potently inhibited by quinidine (CYP2D inhibitor) but had only a minor impact from all CYP inducers tested. Intrinsic clearance estimates showed over 50 times higher values for (+)-M5 formation from (+)-M2 compared with (+)-M1 in DLMs. This was largely attributed to the higher enzyme affinity (lower Km) for (+)-M2 compared with (+)-M1 as substrate. (+)-tramadol, (+)-M1, (+)-M2, or (+)-M5 were not p-glycoprotein substrates. This study provides a clearer picture of the role of individual CYPs in the complex metabolism of tramadol in dogs.
我们之前的研究表明,(+)-曲马多在犬肝脏中仅通过CYP2D15代谢为(+)-M1,通过多种细胞色素P450(CYPs)代谢为(+)-M2,但主要是通过CYP2B11。然而,(+)-M1和(+)-M2在犬体内进一步代谢为(+)-M5,这是在犬血浆和尿液中发现的主要代谢物。在本研究中,我们使用重组酶、未处理的犬肝脏微粒体(DLMs)、抑制剂处理的DLMs以及来自CYP诱导剂处理犬的DLMs,鉴定了参与(+)-M1和(+)-M2代谢的犬CYPs。还使用了一种表达犬P-糖蛋白的细胞系来评估(+)-曲马多、(+)-M1、(+)-M2或(+)-M5是否为犬P-糖蛋白的底物,从而限制它们向中枢神经系统的分布。(+)-M5主要由重组CYP2C21从(+)-M1形成,CYP2C41和CYP2B11的贡献较小。磺胺苯吡唑(CYP2C抑制剂)和氯霉素(CYP2B11抑制剂)强烈抑制了DLMs中(+)-M1形成(+)-M5的过程,而在苯巴比妥处理犬的DLMs中该过程大大增加。(+)-M5主要由CYP2D15从(+)-M2形成。DLMs中(+)-M1形成(+)-M5的过程受到奎尼丁(CYP2D抑制剂)的强烈抑制,但在所有测试的CYP诱导剂作用下影响较小。内在清除率估计显示,DLMs中(+)-M2形成(+)-M5的值比(+)-M1形成(+)-M5的值高50倍以上。这主要归因于与作为底物的(+)-M1相比,(+)-M2具有更高的酶亲和力(更低的Km)。(+)-曲马多、(+)-M1、(+)-M2或(+)-M5不是P-糖蛋白底物。本研究更清晰地描绘了个体CYPs在犬曲马多复杂代谢中的作用。