Center for Molecular Biology of Heidelberg University (ZMBH), DKFZ-ZMBH Alliance, Heidelberg, Germany.
German Cancer Research Center (DKFZ), Heidelberg, Germany.
Nature. 2018 Sep;561(7722):268-272. doi: 10.1038/s41586-018-0462-y. Epub 2018 Aug 29.
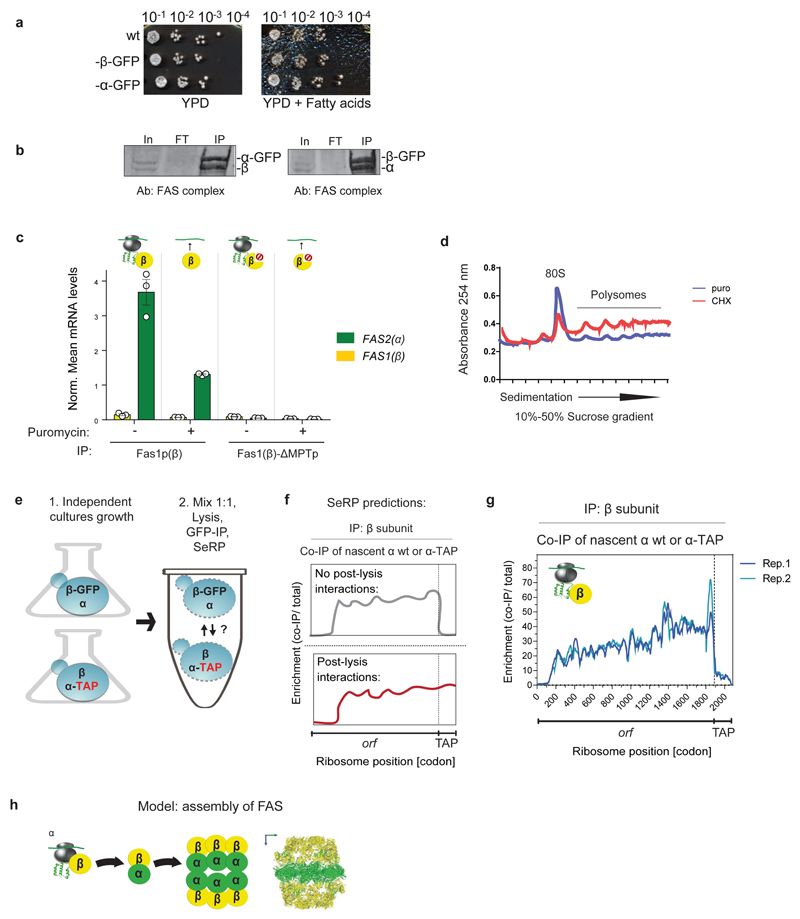
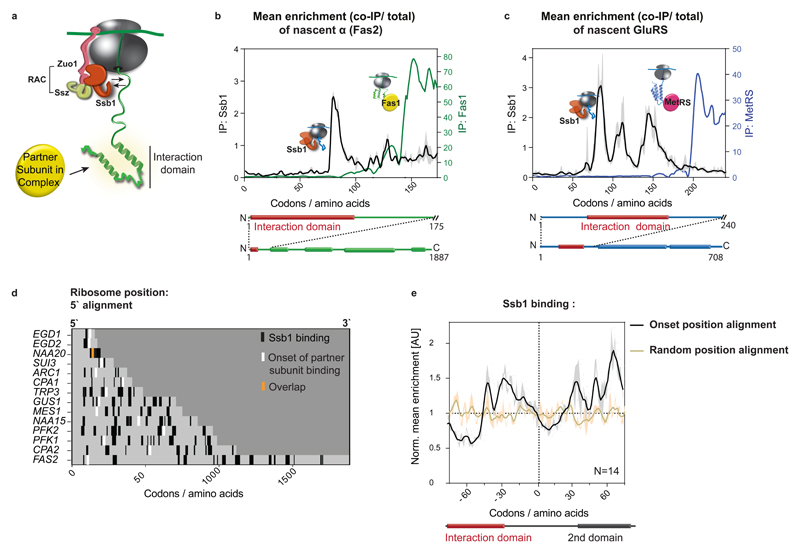
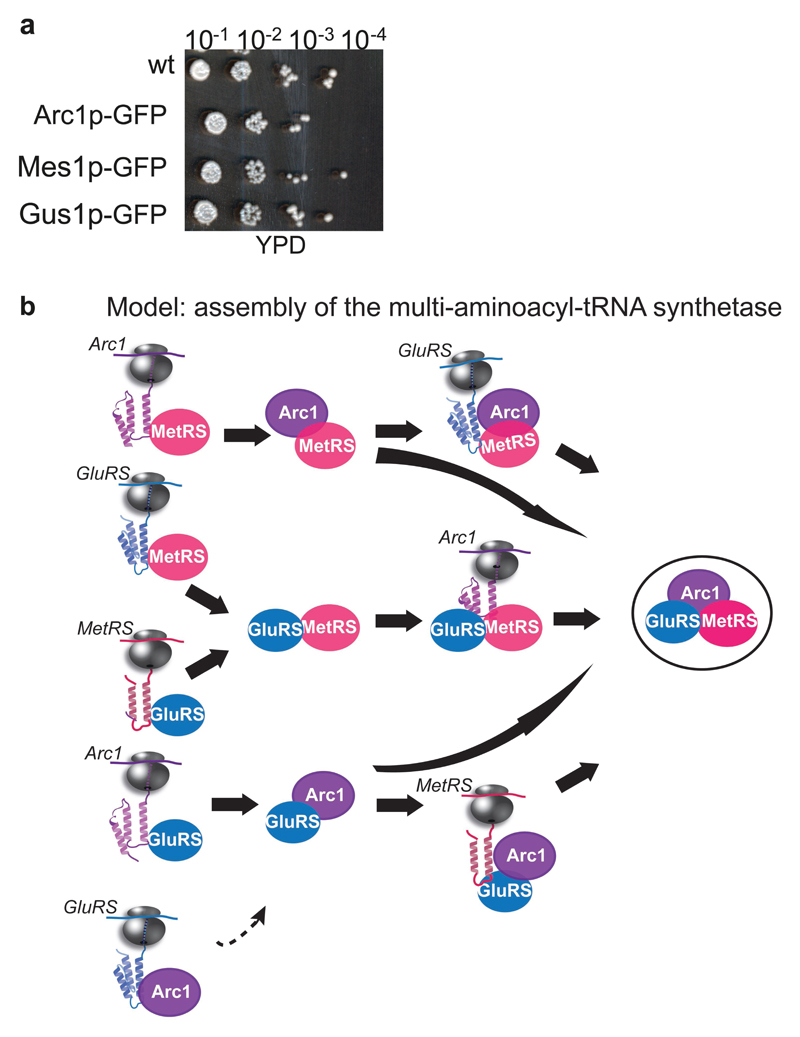
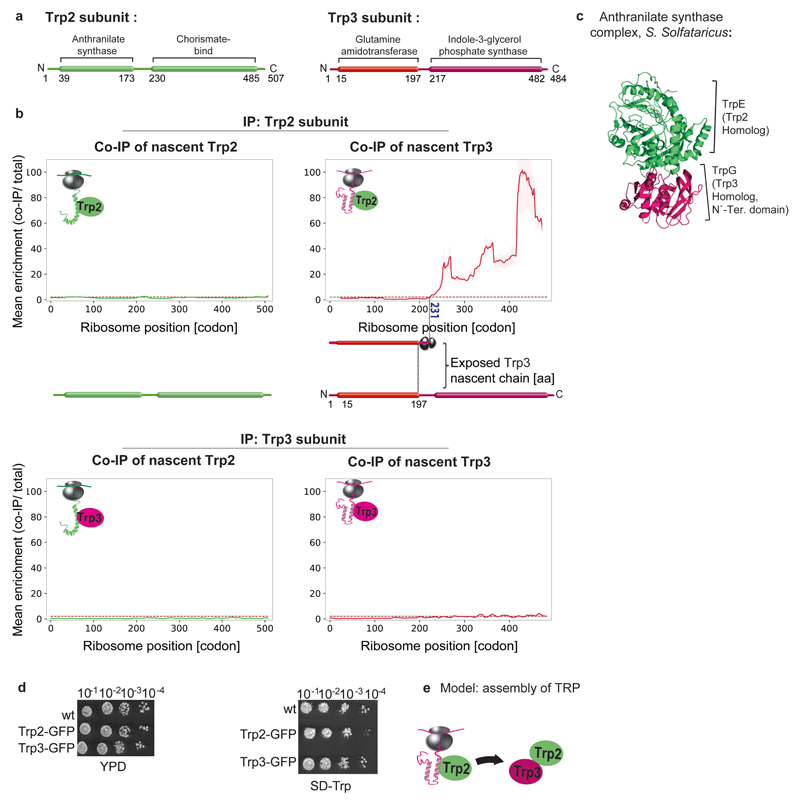
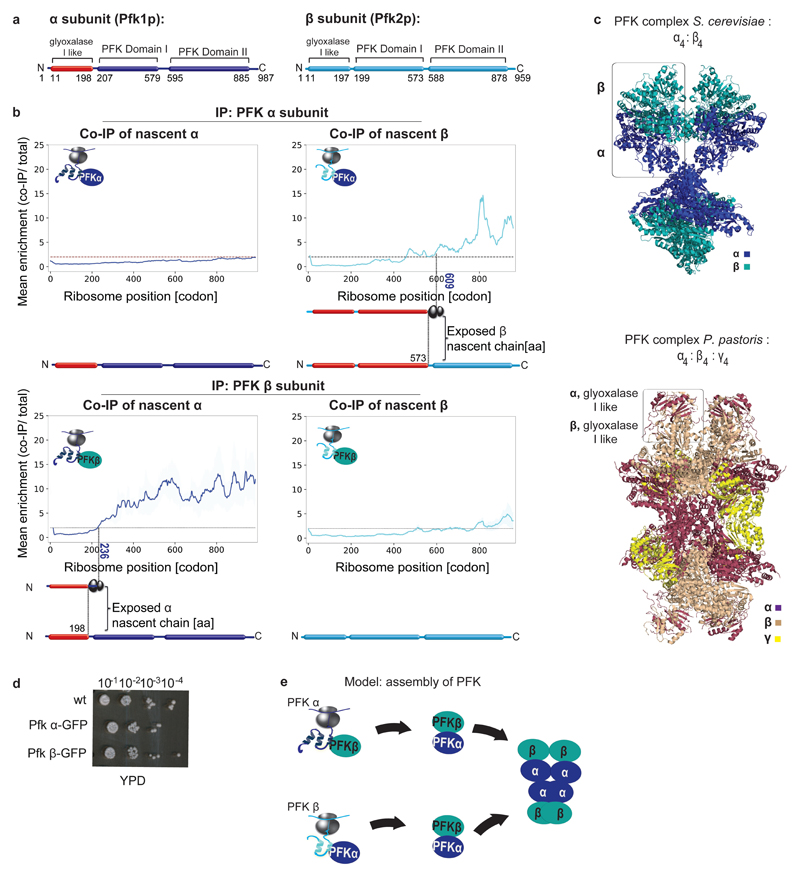
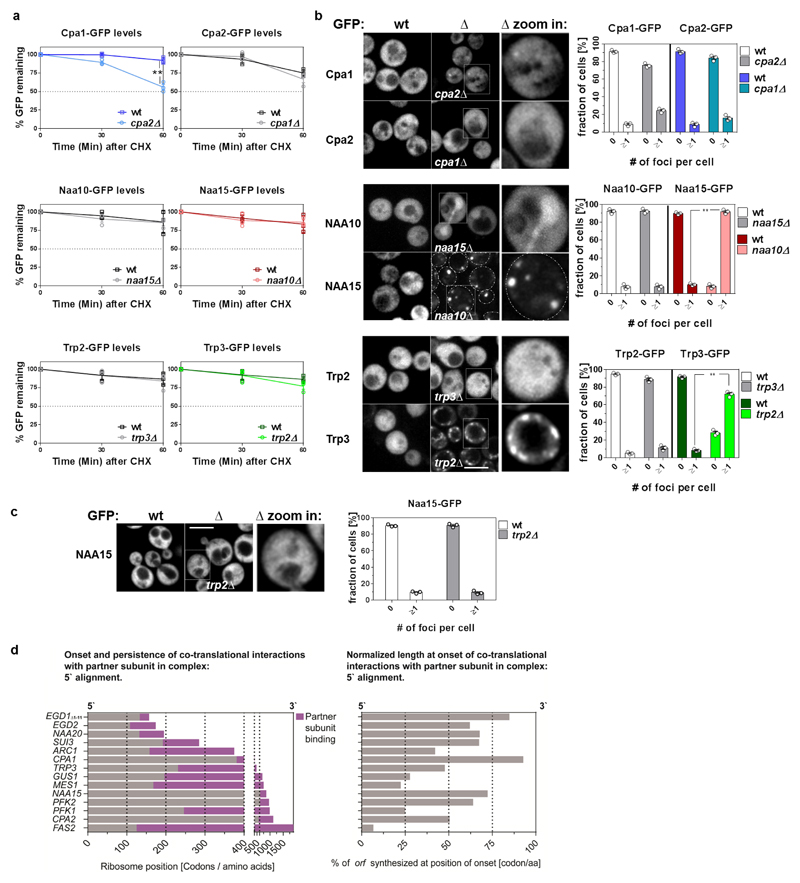
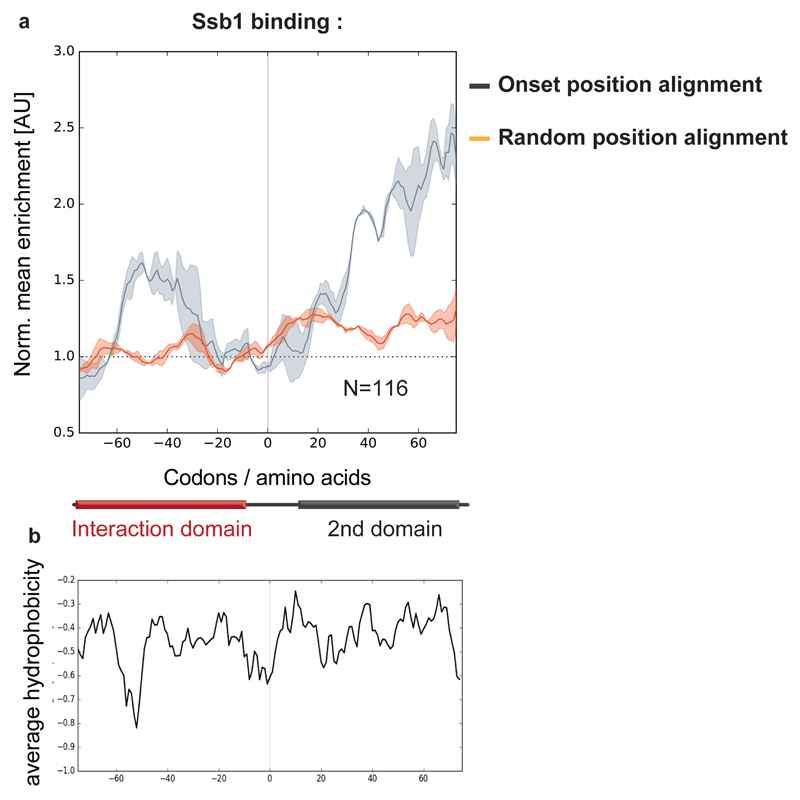
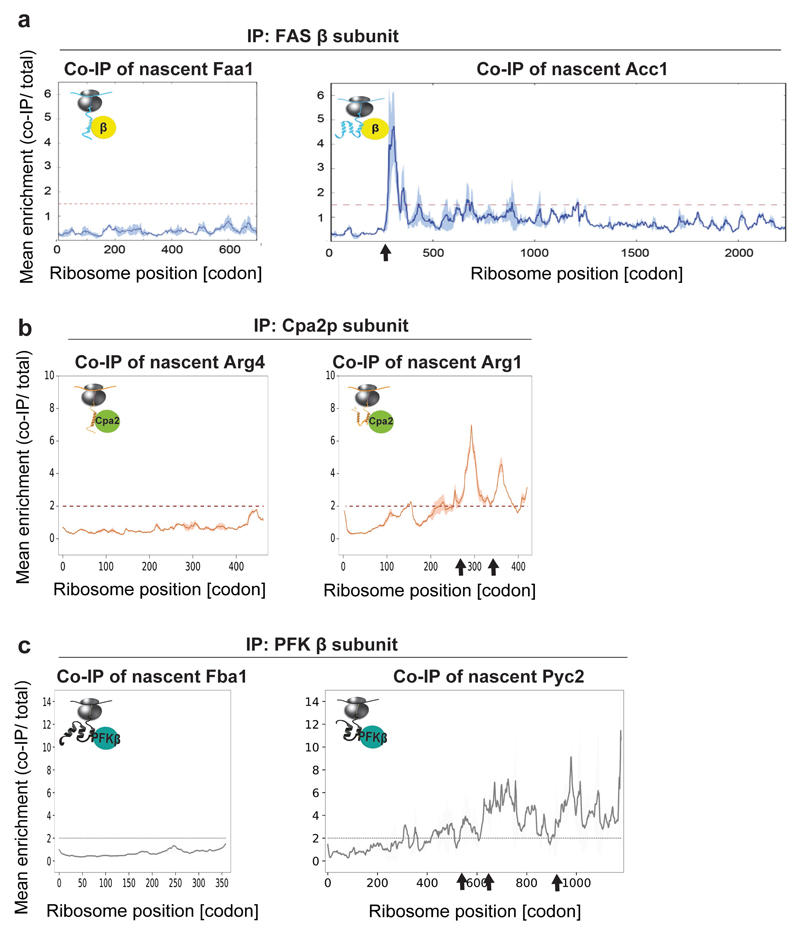
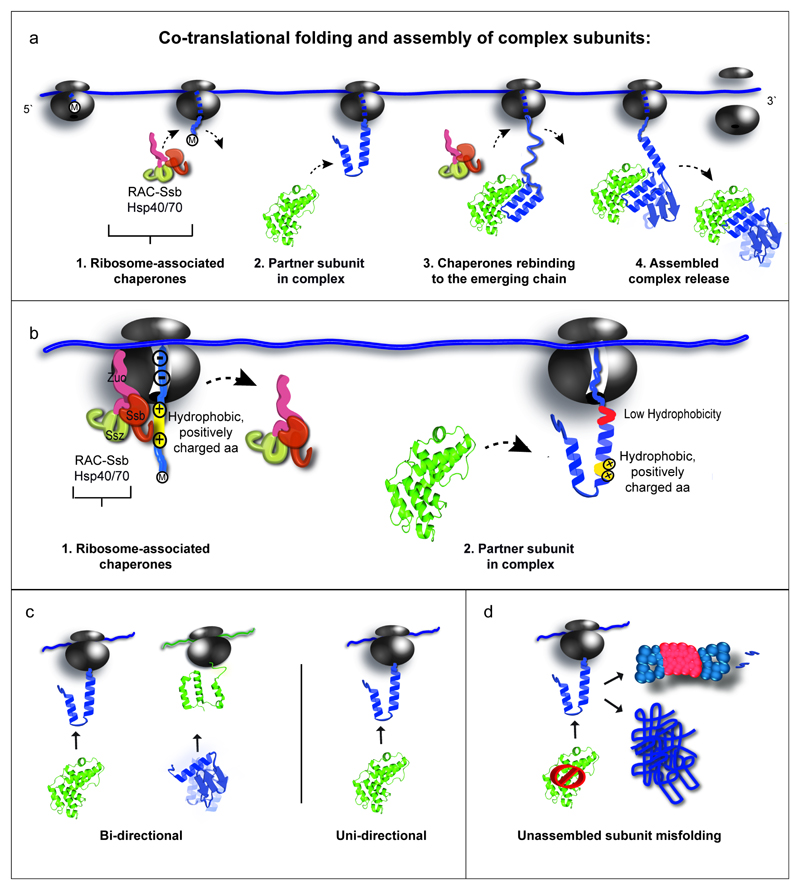
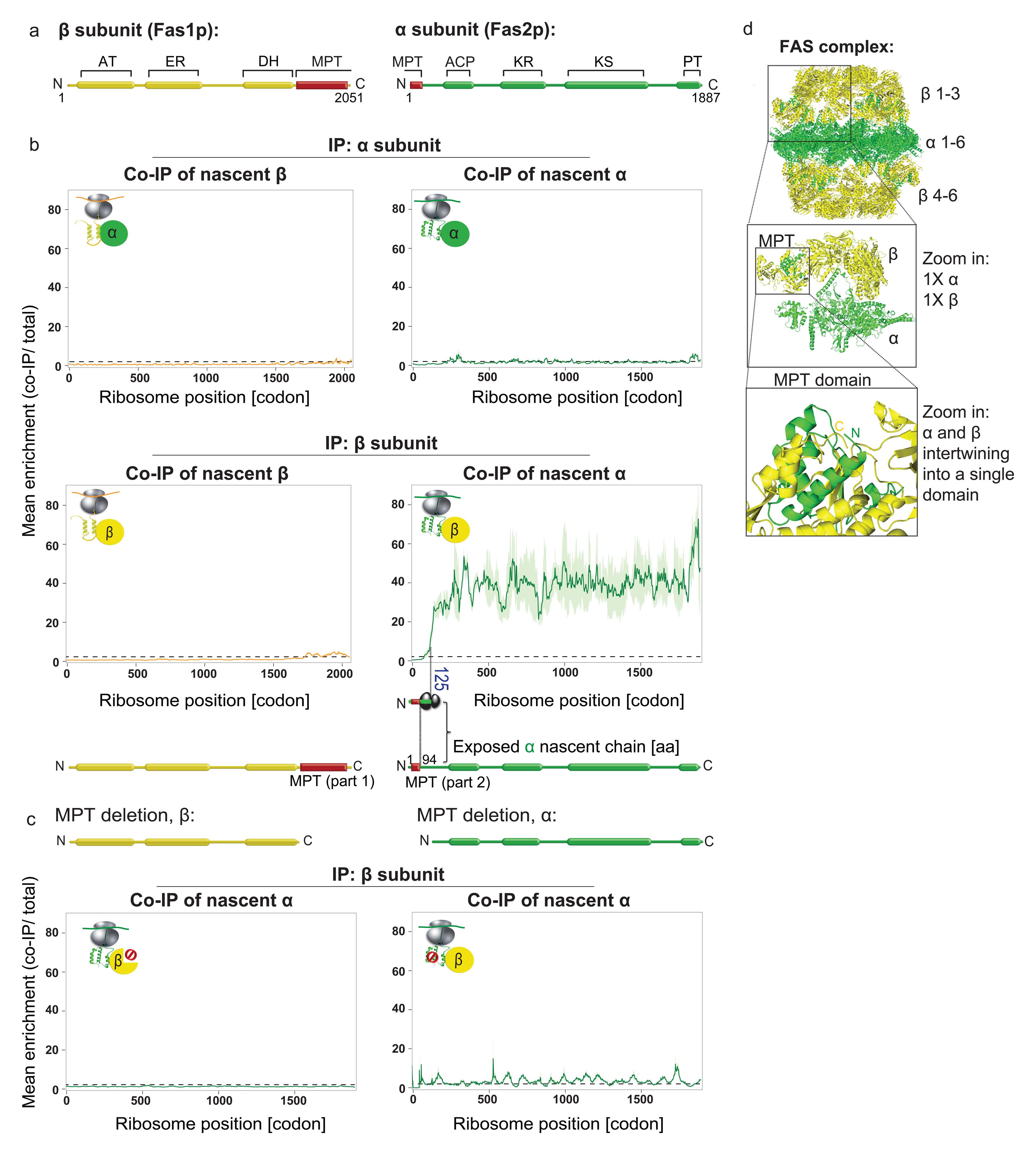
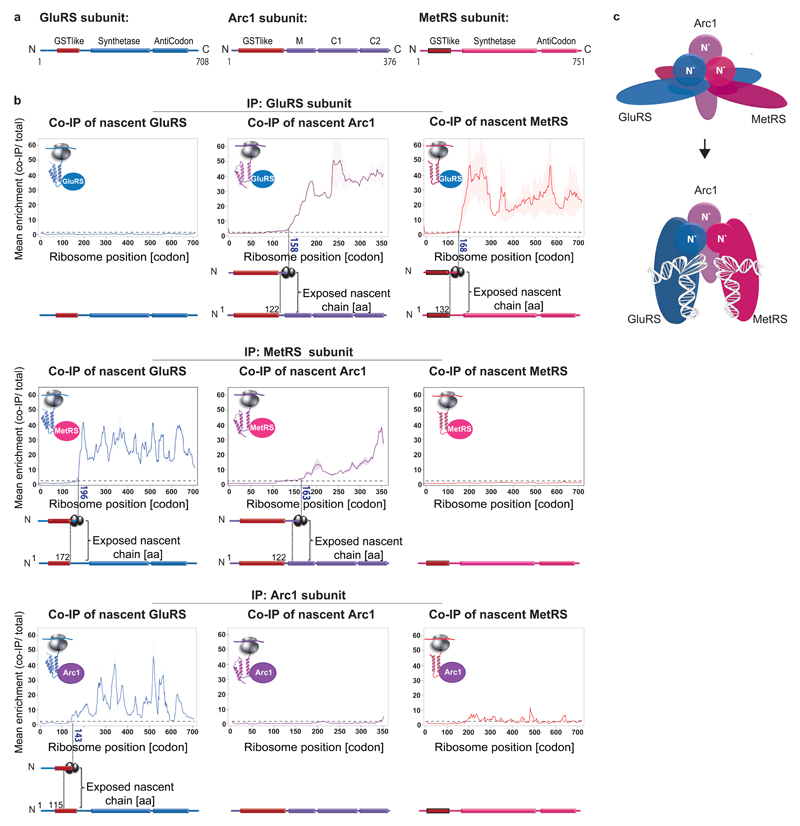
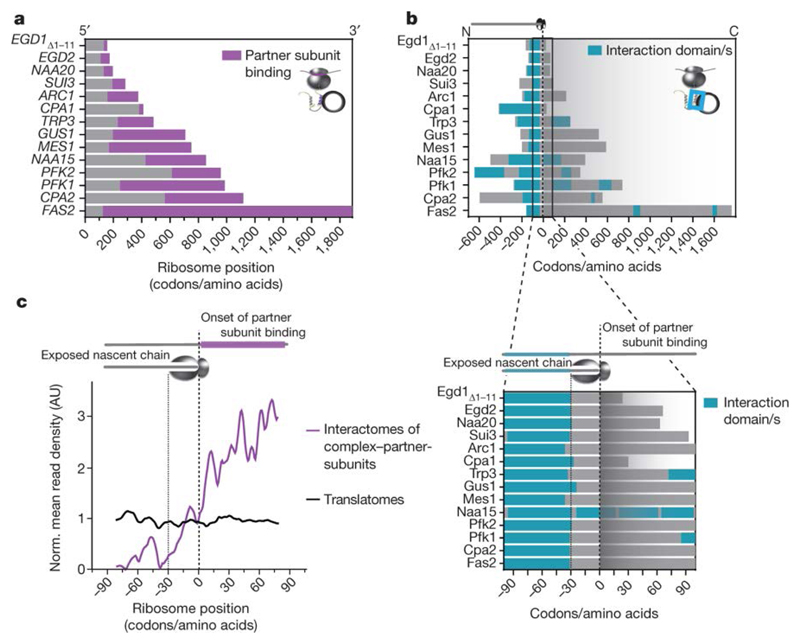
The folding of newly synthesized proteins to the native state is a major challenge within the crowded cellular environment, as non-productive interactions can lead to misfolding, aggregation and degradation. Cells cope with this challenge by coupling synthesis with polypeptide folding and by using molecular chaperones to safeguard folding cotranslationally. However, although most of the cellular proteome forms oligomeric assemblies, little is known about the final step of folding: the assembly of polypeptides into complexes. In prokaryotes, a proof-of-concept study showed that the assembly of heterodimeric luciferase is an organized cotranslational process that is facilitated by spatially confined translation of the subunits encoded on a polycistronic mRNA. In eukaryotes, however, fundamental differences-such as the rarity of polycistronic mRNAs and different chaperone constellations-raise the question of whether assembly is also coordinated with translation. Here we provide a systematic and mechanistic analysis of the assembly of protein complexes in eukaryotes using ribosome profiling. We determined the in vivo interactions of the nascent subunits from twelve hetero-oligomeric protein complexes of Saccharomyces cerevisiae at near-residue resolution. We find nine complexes assemble cotranslationally; the three complexes that do not show cotranslational interactions are regulated by dedicated assembly chaperones. Cotranslational assembly often occurs uni-directionally, with one fully synthesized subunit engaging its nascent partner subunit, thereby counteracting its propensity for aggregation. The onset of cotranslational subunit association coincides directly with the full exposure of the nascent interaction domain at the ribosomal tunnel exit. The action of the ribosome-associated Hsp70 chaperone Ssb is coordinated with assembly. Ssb transiently engages partially synthesized interaction domains and then dissociates before the onset of partner subunit association, presumably to prevent premature assembly interactions. Our study shows that cotranslational subunit association is a prevalent mechanism for the assembly of hetero-oligomers in yeast and indicates that translation, folding and the assembly of protein complexes are integrated processes in eukaryotes.
新合成的蛋白质折叠成天然状态是细胞环境中面临的主要挑战,因为非生产性相互作用会导致错误折叠、聚集和降解。细胞通过将合成与多肽折叠偶联,并利用分子伴侣在共翻译过程中保护折叠来应对这一挑战。然而,尽管细胞中的大部分蛋白质组形成寡聚体组装,但对于折叠的最后一步:多肽组装成复合物,我们知之甚少。在原核生物中,一项概念验证研究表明,异二聚体荧光素酶的组装是一个有组织的共翻译过程,该过程由在多顺反子 mRNA 上编码的亚基的空间受限翻译来促进。然而,在真核生物中,由于多顺反子 mRNA 的罕见性和不同的伴侣蛋白组合等基本差异,引发了一个问题,即组装是否也与翻译协调。在这里,我们使用核糖体谱分析对真核生物中蛋白质复合物的组装进行了系统和机制分析。我们在活体内确定了来自酿酒酵母的十二个异源寡聚体蛋白复合物的新生亚基的相互作用,分辨率接近残基水平。我们发现有九个复合物进行共翻译组装;另外三个没有表现出共翻译相互作用的复合物由专门的组装伴侣蛋白调节。共翻译组装通常是单向进行的,一个完全合成的亚基与它的新生伴侣亚基结合,从而抵消了它聚集的倾向。共翻译亚基结合的开始与新生相互作用结构域在核糖体隧道出口处完全暴露直接吻合。核糖体相关 Hsp70 伴侣 Ssb 的作用与组装相协调。Ssb 短暂地与部分合成的相互作用结构域结合,然后在伴侣亚基结合之前解离,大概是为了防止过早的组装相互作用。我们的研究表明,共翻译亚基结合是酵母中异源寡聚体组装的一种常见机制,并表明翻译、折叠和蛋白质复合物的组装是真核生物中整合的过程。