Brodmann Maj, Heilig Rosalie, Broz Petr, Basler Marek
Biozentrum, University of Basel, Basel, Switzerland.
Department of Biochemistry, University of Lausanne, Épalinges, Switzerland.
Front Cell Infect Microbiol. 2018 Aug 31;8:284. doi: 10.3389/fcimb.2018.00284. eCollection 2018.
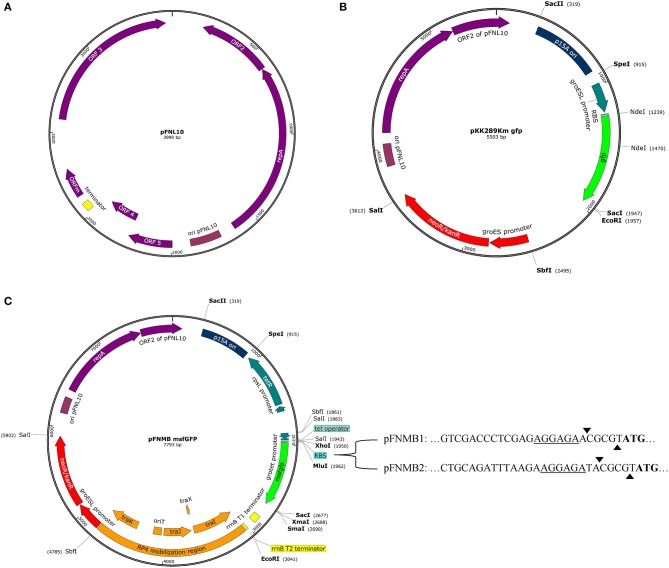
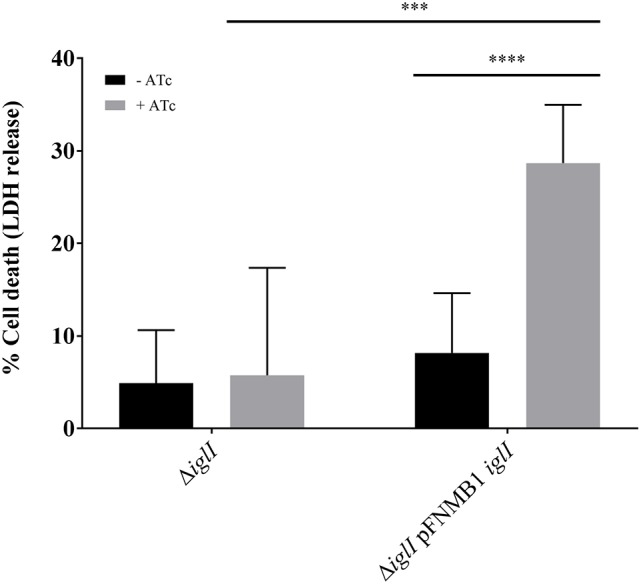
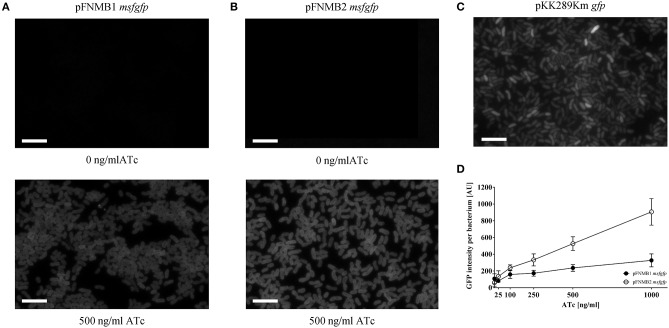
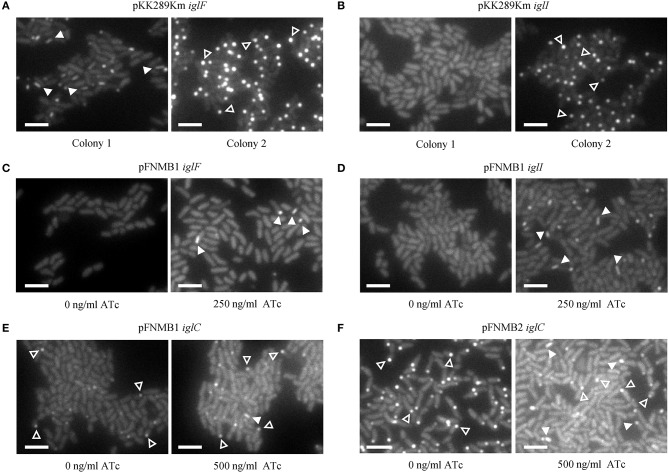
is the causative agent of the life-threatening disease tularemia. However, the molecular tools to study are limited. Especially, expression plasmids are sparse and difficult to use, as they are unstable and prone to spontaneous loss. Most expression plasmids lack inducible promoters making it difficult to control gene expression levels. In addition, available expression plasmids are mainly designed for , however, genetic differences including restriction-modification systems impede the use of these plasmids in , which is often used as a model organism to study pathogenesis. Here we report construction and characterization of two mobilizable plasmids (pFNMB1 and pFNMB2) designed for regulated gene expression in . pFNMB plasmids contain a tetracycline inducible promoter to control gene expression levels and for RP4 mediated mobilization. We show that both plasmids are stably maintained in bacteria for more than 40 generations over 4 days of culturing in the absence of selection against plasmid loss. Expression levels are dependent on anhydrotetracycline concentration and homogeneous in a bacterial population. pFNMB1 and pFNMB2 plasmids differ in the sequence between promoter and translation start site and thus allow to reach different maximum levels of protein expression. We used pFNMB1 and pFNMB2 for complementation of Pathogenicity Island mutants Δ, Δ, and Δ and pFNMB1 to complement Δ mutant in bone marrow derived macrophages.
是危及生命的疾病兔热病的病原体。然而,用于研究的分子工具有限。特别是,表达质粒稀少且难以使用,因为它们不稳定且容易自发丢失。大多数表达质粒缺乏诱导型启动子,使得难以控制基因表达水平。此外,可用的表达质粒主要是为设计的,然而,包括限制修饰系统在内的遗传差异阻碍了这些质粒在中的使用,而通常被用作研究发病机制的模式生物。在此,我们报告了两种可移动质粒(pFNMB1和pFNMB2)的构建和特性,这些质粒设计用于在中进行调控基因表达。pFNMB质粒含有四环素诱导型启动子以控制基因表达水平,并具有用于RP4介导的转移的元件。我们表明,在没有针对质粒丢失的选择的情况下,两种质粒在细菌中培养4天超过40代仍能稳定维持。表达水平取决于脱水四环素的浓度且在细菌群体中是均匀的。pFNMB1和pFNMB2质粒在启动子和翻译起始位点之间的序列不同,因此能够达到不同的最大蛋白质表达水平。我们使用pFNMB1和pFNMB2对致病性岛突变体Δ、Δ和Δ进行互补,并使用pFNMB1对骨髓来源巨噬细胞中的Δ突变体进行互补。