Department of Botany, University of British Columbia, 6270 University Blvd, Vancouver, BC, V6T 1Z4, Canada.
Division of Invertebrate Zoology and Sackler Institute for Comparative Genomics, American Museum of Natural History, 200 Central Park West, New York, NY, 10024, USA.
BMC Biol. 2018 Nov 5;16(1):136. doi: 10.1186/s12915-018-0605-5.
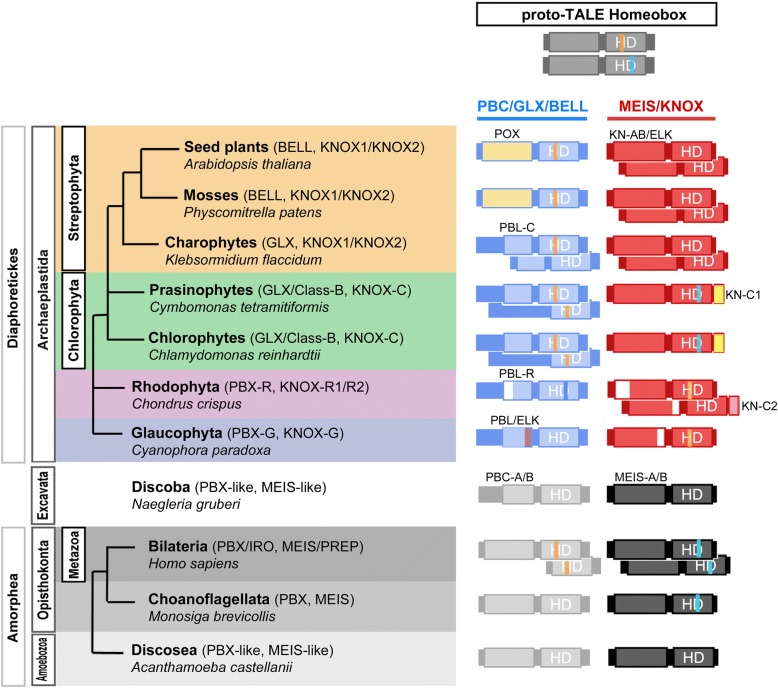
Complex multicellularity requires elaborate developmental mechanisms, often based on the versatility of heterodimeric transcription factor (TF) interactions. Homeobox TFs in the TALE superclass are deeply embedded in the gene regulatory networks that orchestrate embryogenesis. Knotted-like homeobox (KNOX) TFs, homologous to animal MEIS, have been found to drive the haploid-to-diploid transition in both unicellular green algae and land plants via heterodimerization with other TALE superclass TFs, demonstrating remarkable functional conservation of a developmental TF across lineages that diverged one billion years ago. Here, we sought to delineate whether TALE-TALE heterodimerization is ancestral to eukaryotes.
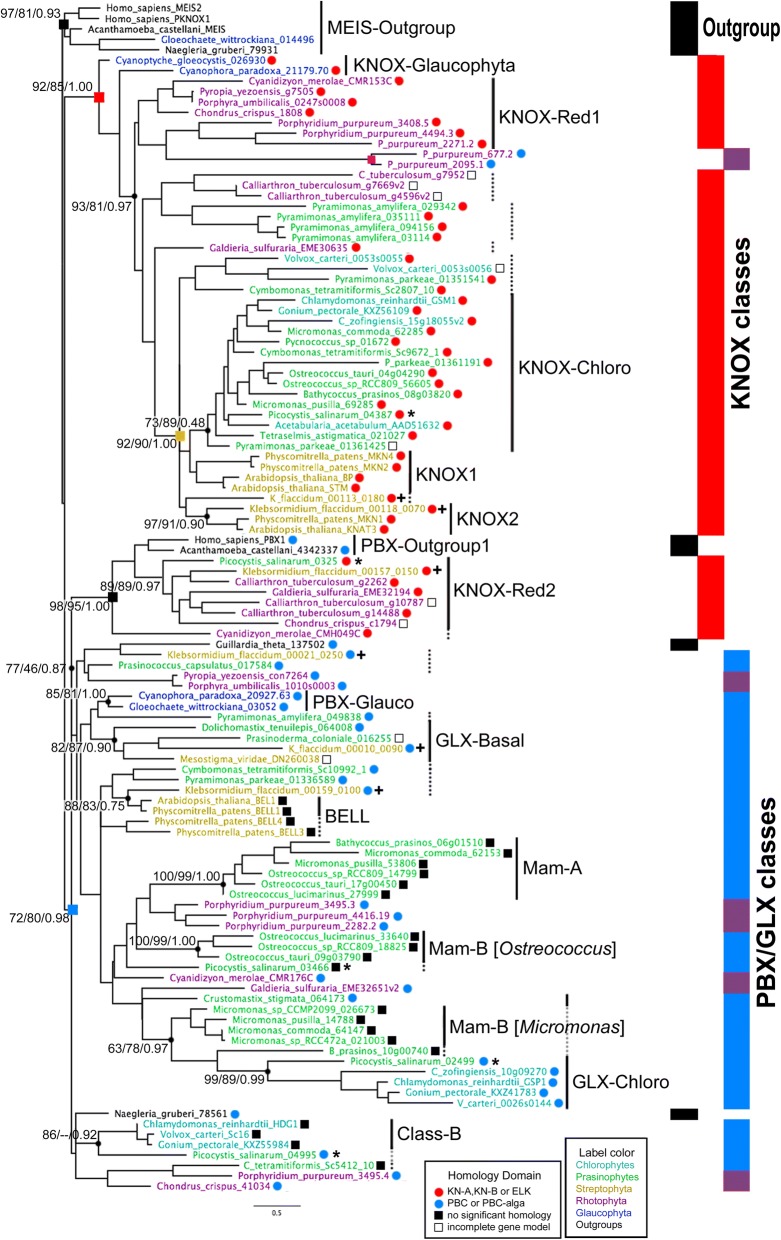
We analyzed TALE endowment in the algal radiations of Archaeplastida, ancestral to land plants. Homeodomain phylogeny and bioinformatics analysis partitioned TALEs into two broad groups, KNOX and non-KNOX. Each group shares previously defined heterodimerization domains, plant KNOX-homology in the KNOX group and animal PBC-homology in the non-KNOX group, indicating their deep ancestry. Protein-protein interaction experiments showed that the TALEs in the two groups all participated in heterodimerization.
Our study indicates that the TF dyads consisting of KNOX/MEIS and PBC-containing TALEs must have evolved early in eukaryotic evolution. Based on our results, we hypothesize that in early eukaryotes, the TALE heterodimeric configuration provided transcription-on switches via dimerization-dependent subcellular localization, ensuring execution of the haploid-to-diploid transition only when the gamete fusion is correctly executed between appropriate partner gametes. The TALE switch then diversified in the several lineages that engage in a complex multicellular organization.
复杂的多细胞性需要精细的发育机制,这些机制通常基于异二聚体转录因子 (TF) 相互作用的多功能性。TALE 超家族中的同源盒 TF 深深嵌入协调胚胎发生的基因调控网络中。与动物 MEIS 同源的类 Knotted 同源盒 (KNOX) TF 已被发现通过与其他 TALE 超家族 TF 异二聚化,在单细胞绿藻和陆地植物中驱动从单倍体到二倍体的转变,证明了一个发育 TF 在 10 亿年前分化的谱系中具有显著的功能保守性。在这里,我们试图描绘 TALE-TALE 异二聚化是否是真核生物的祖先。
我们分析了 Archaeplastida 藻类辐射中的 TALE 赋存情况,它是陆地植物的祖先。同源盒系统发育和生物信息学分析将 TALEs 分为两大组,KNOX 和非-KNOX。每组都共享先前定义的异二聚化结构域,KNOX 组中的 KNOX 同源结构域和非-KNOX 组中的动物 PBC 同源结构域,表明它们的深度祖先。蛋白质-蛋白质相互作用实验表明,这两组中的 TALEs 都参与了异二聚化。
我们的研究表明,由 KNOX/MEIS 和包含 PBC 的 TALEs 组成的 TF 二聚体必须在真核生物进化的早期就已经进化。基于我们的结果,我们假设在早期的真核生物中,TALE 异二聚体构型通过依赖于二聚体的亚细胞定位的二聚化提供了转录开启开关,从而确保只有在适当的配子之间正确执行配子融合时,才会执行从单倍体到二倍体的转变。然后,TALE 开关在参与复杂多细胞组织的几个谱系中多样化。