Majerska Jana, Feretzaki Marianna, Glousker Galina, Lingner Joachim
School of Life Sciences, Ecole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland.
Swiss Institute for Experimental Cancer Research, Ecole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland.
Life Sci Alliance. 2018 Jul 17;1(4):e201800121. doi: 10.26508/lsa.201800121. eCollection 2018 Aug.
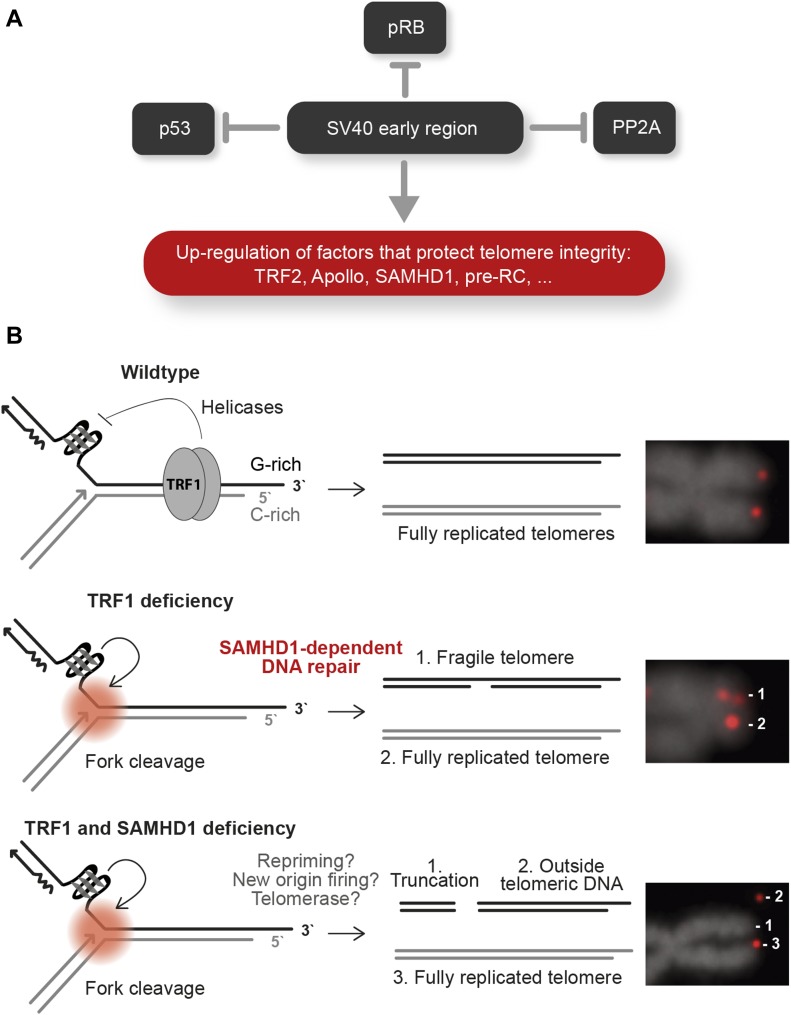
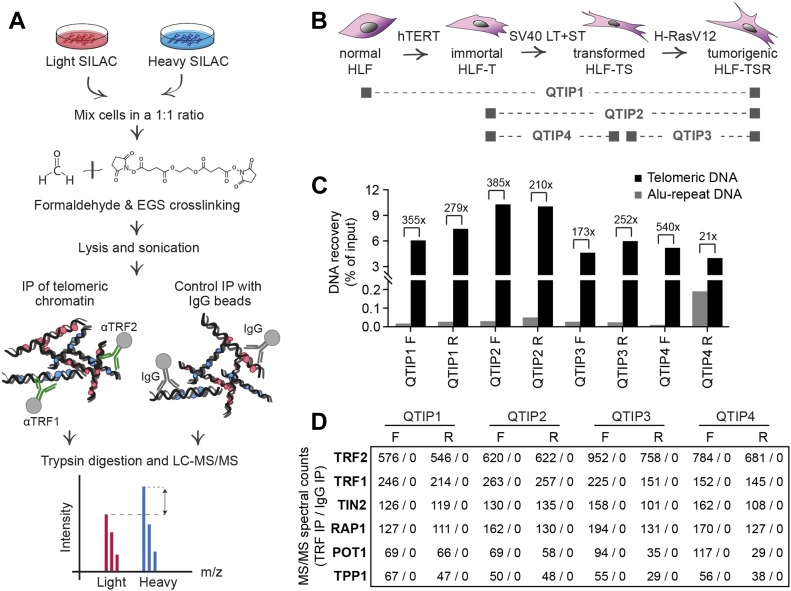
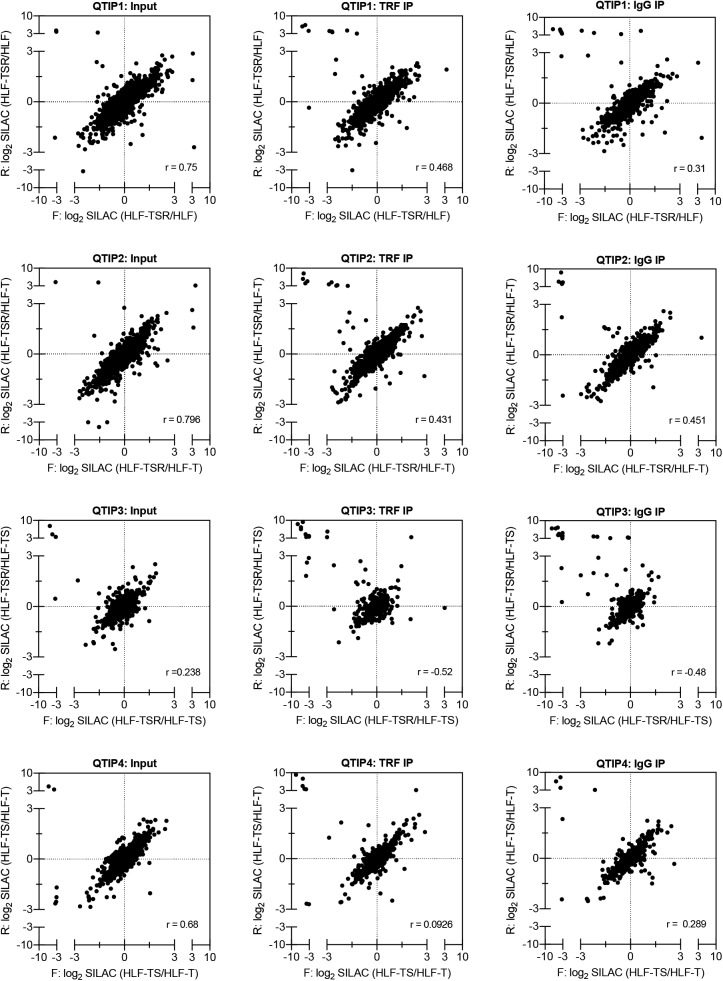
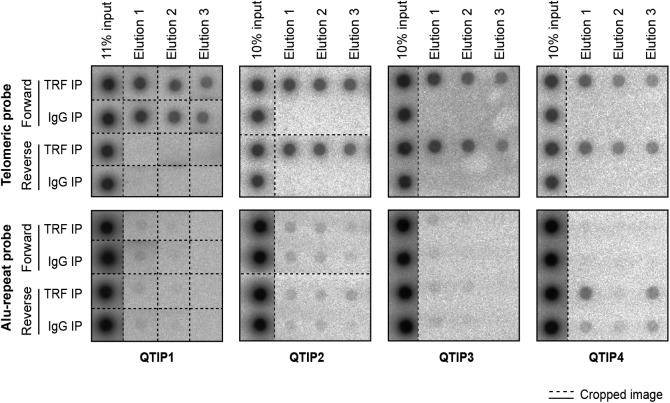
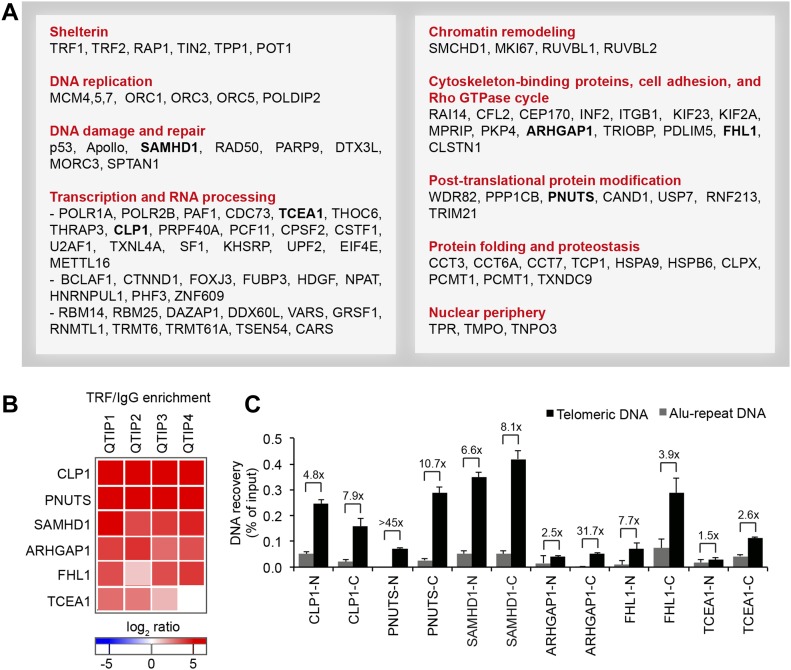
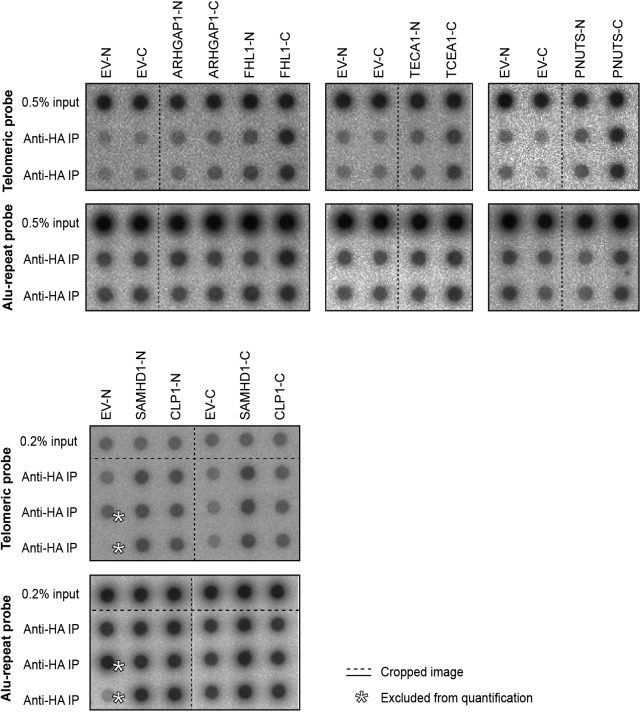
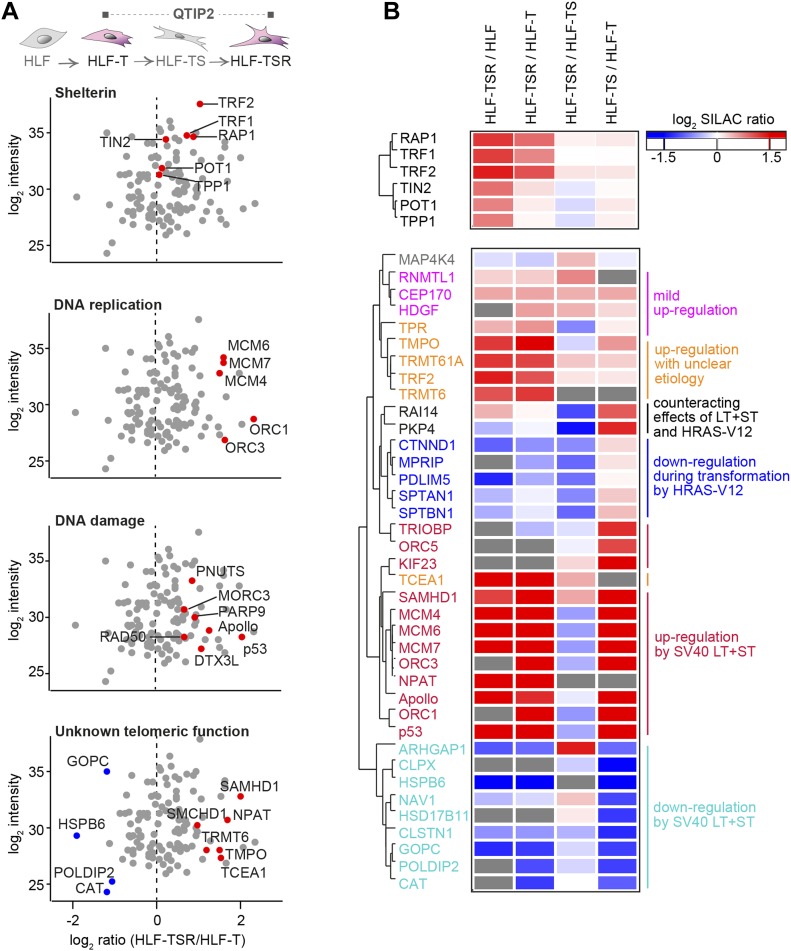
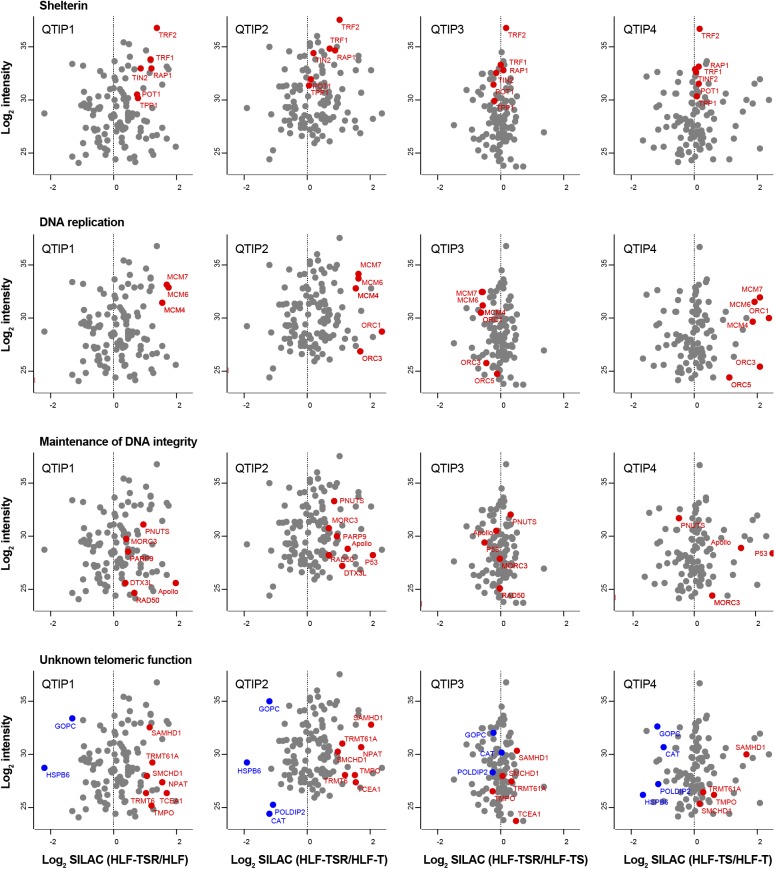
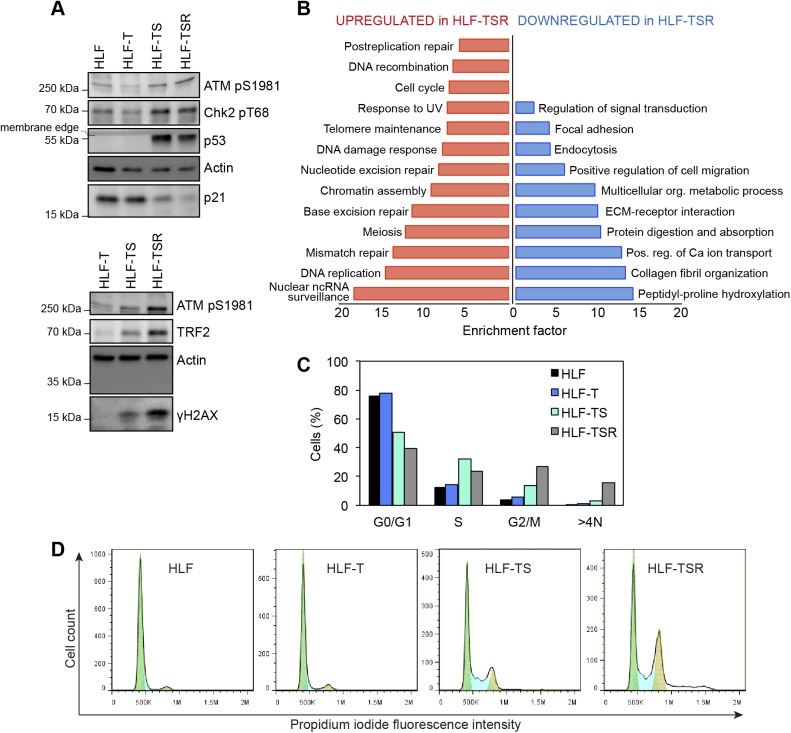
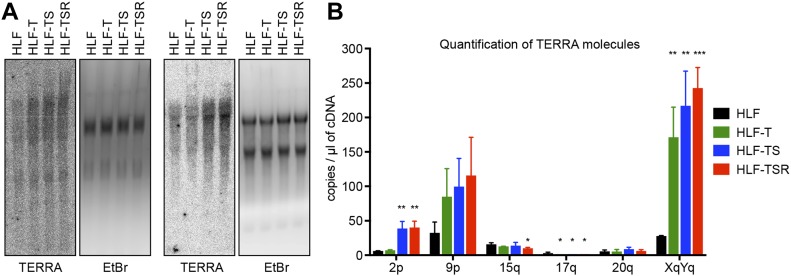
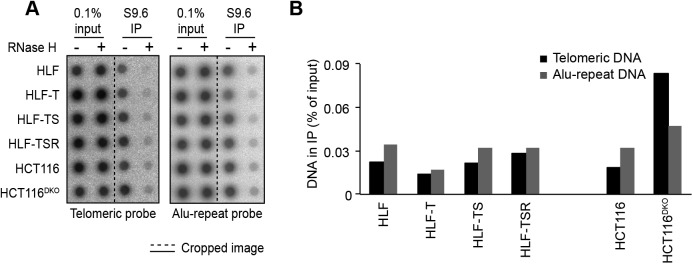
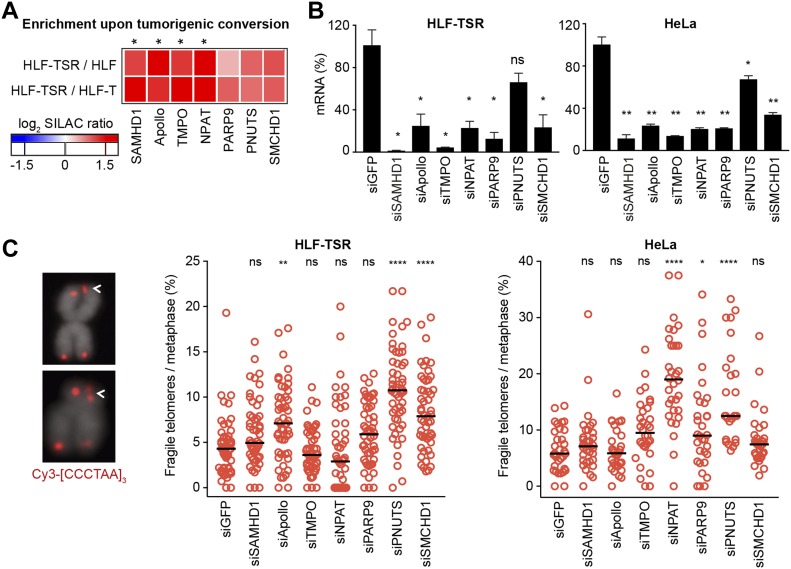
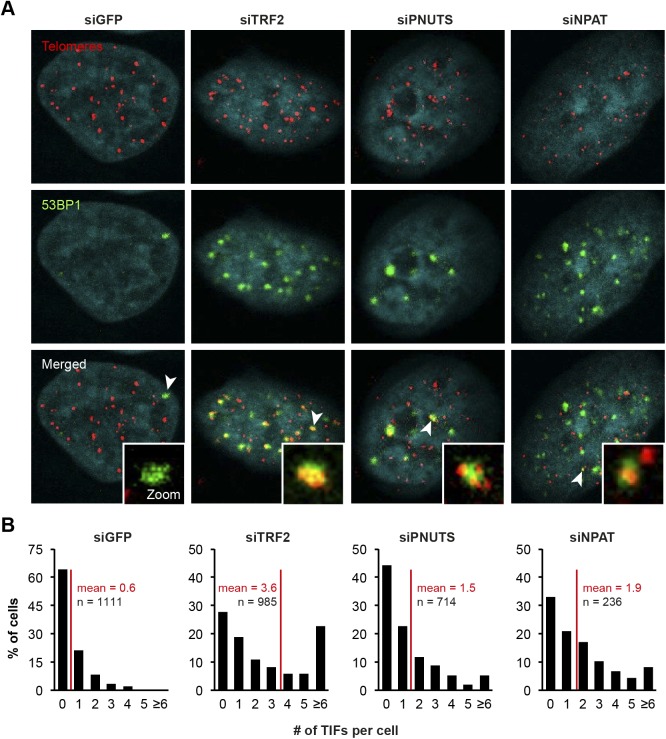
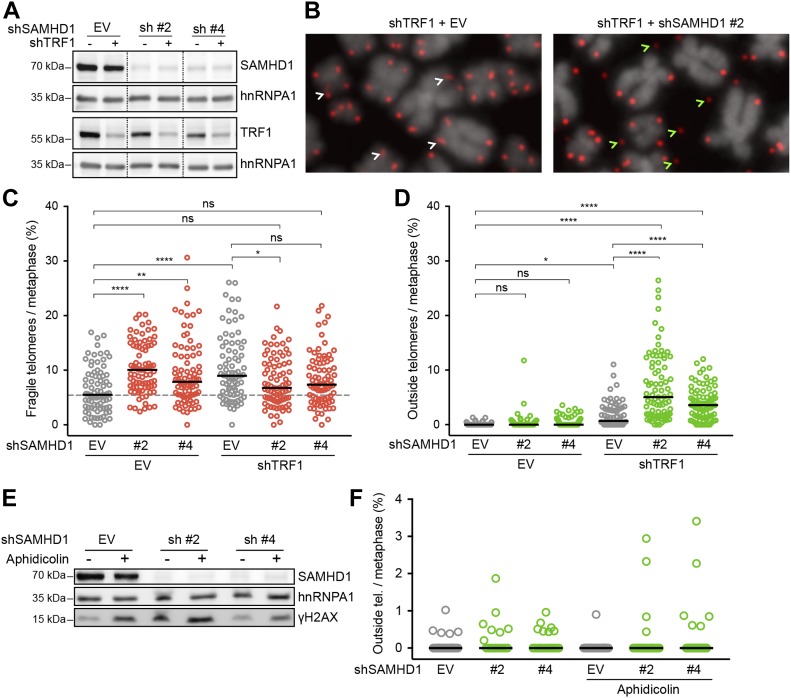
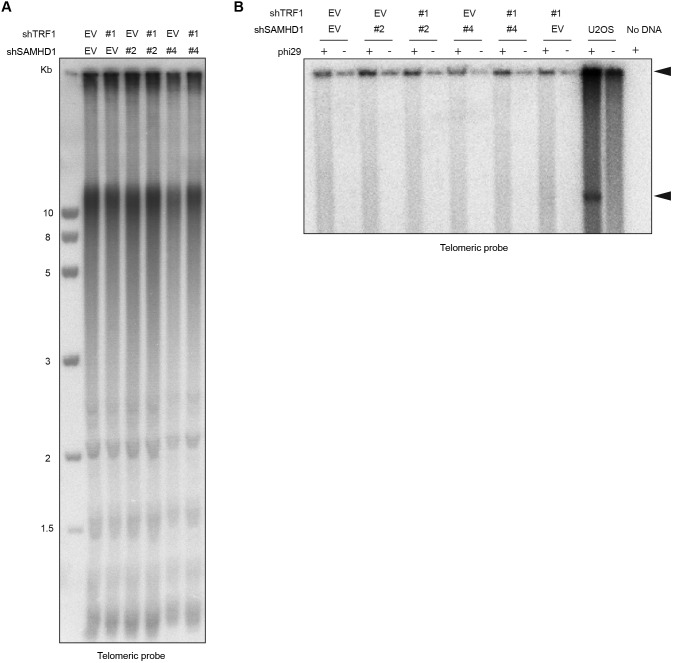
Telomeres play crucial roles during tumorigenesis, inducing cellular senescence upon telomere shortening and extensive chromosome instability during telomere crisis. However, it has not been investigated if and how cellular transformation and oncogenic stress alter telomeric chromatin composition and function. Here, we transform human fibroblasts by consecutive transduction with vectors expressing hTERT, the SV40 early region, and activated H-RasV12. Pairwise comparisons of the telomeric proteome during different stages of transformation reveal up-regulation of proteins involved in chromatin remodeling, DNA repair, and replication at chromosome ends. Depletion of several of these proteins induces telomere fragility, indicating their roles in replication of telomeric DNA. Depletion of SAMHD1, which has reported roles in DNA resection and homology-directed repair, leads to telomere breakage events in cells deprived of the shelterin component TRF1. Thus, our analysis identifies factors, which accumulate at telomeres during cellular transformation to promote telomere replication and repair, resisting oncogene-borne telomere replication stress.
端粒在肿瘤发生过程中发挥着关键作用,在端粒缩短时诱导细胞衰老,并在端粒危机期间引发广泛的染色体不稳定。然而,细胞转化和致癌应激是否以及如何改变端粒染色质组成和功能尚未得到研究。在这里,我们通过连续转导表达hTERT、SV40早期区域和激活的H-RasV12的载体来转化人成纤维细胞。在转化的不同阶段对端粒蛋白质组进行成对比较,发现参与染色质重塑、DNA修复和染色体末端复制的蛋白质上调。其中几种蛋白质的缺失会诱导端粒脆性,表明它们在端粒DNA复制中的作用。SAMHD1在DNA切除和同源定向修复中发挥作用,其缺失会导致缺乏保护蛋白组分TRF1的细胞中发生端粒断裂事件。因此,我们的分析确定了在细胞转化过程中积累在端粒上以促进端粒复制和修复、抵抗癌基因引起的端粒复制应激的因素。