Department of Medicine, Division of Hematology, University of Washington, Seattle, WA; and.
Institute for Systems Biology, Seattle, WA.
Blood. 2019 Jan 31;133(5):457-469. doi: 10.1182/blood-2018-05-850412. Epub 2018 Dec 10.
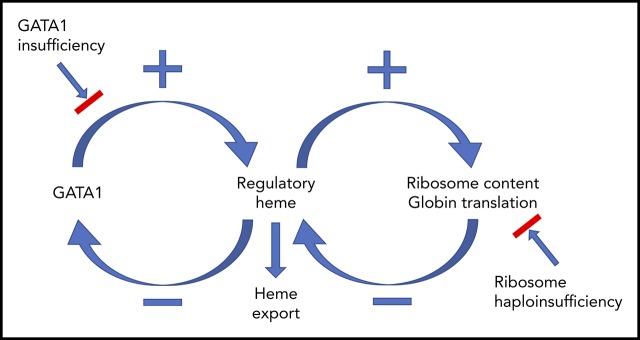
Erythropoiesis is the complex, dynamic, and tightly regulated process that generates all mature red blood cells. To understand this process, we mapped the developmental trajectories of progenitors from wild-type, erythropoietin-treated, and -deleted mice at single-cell resolution. Importantly, we linked the quantity of each cell's surface proteins to its total transcriptome, which is a novel method. Deletion of results in high levels of intracellular heme, allowing us to identify heme-regulated circuitry. Our studies demonstrate that in early erythroid cells (CD71Ter119), heme increases ribosomal protein transcripts, suggesting that heme, in addition to upregulating globin transcription and translation, guarantees ample ribosomes for globin synthesis. In later erythroid cells (CD71Ter119), heme decreases GATA1, GATA1-target gene, and mitotic spindle gene expression. These changes occur quickly. For example, in confirmatory studies using human marrow erythroid cells, ribosomal protein transcripts and proteins increase, and GATA1 transcript and protein decrease, within 15 to 30 minutes of amplifying endogenous heme synthesis with aminolevulinic acid. Because GATA1 initiates heme synthesis, GATA1 and heme together direct red cell maturation, and heme stops GATA1 synthesis, our observations reveal a GATA1-heme autoregulatory loop and implicate GATA1 and heme as the comaster regulators of the normal erythroid differentiation program. In addition, as excessive heme could amplify ribosomal protein imbalance, prematurely lower GATA1, and impede mitosis, these data may help explain the ineffective (early termination of) erythropoiesis in Diamond Blackfan anemia and del(5q) myelodysplasia, disorders with excessive heme in colony-forming unit-erythroid/proerythroblasts, explain why these anemias are macrocytic, and show why children with GATA1 mutations have DBA-like clinical phenotypes.
红细胞生成是一个复杂、动态和严格调控的过程,它产生所有成熟的红细胞。为了理解这个过程,我们以单细胞分辨率绘制了来自野生型、促红细胞生成素处理和 - 缺失小鼠的祖细胞的发育轨迹。重要的是,我们将每个细胞表面蛋白的数量与其总转录组联系起来,这是一种新方法。缺失导致细胞内血红素水平升高,使我们能够识别血红素调节回路。我们的研究表明,在早期红细胞(CD71Ter119)中,血红素增加核糖体蛋白转录物,表明血红素除了上调珠蛋白转录和翻译外,还为珠蛋白合成提供充足的核糖体。在晚期红细胞(CD71Ter119)中,血红素降低 GATA1、GATA1 靶基因和有丝分裂纺锤体基因的表达。这些变化发生得很快。例如,在使用人骨髓红细胞进行的验证研究中,在用氨基酮戊酸扩增内源性血红素合成后 15 至 30 分钟内,核糖体蛋白转录物和蛋白质增加,而 GATA1 转录物和蛋白质减少。因为 GATA1 启动血红素合成,GATA1 和血红素一起指导红细胞成熟,血红素停止 GATA1 合成,我们的观察结果揭示了一个 GATA1-血红素自调节回路,并暗示 GATA1 和血红素是正常红细胞分化程序的共同主调节因子。此外,由于过多的血红素可能会放大核糖体蛋白失衡,过早降低 GATA1,并阻碍有丝分裂,这些数据可能有助于解释 Diamond Blackfan 贫血和 del(5q)骨髓增生异常综合征中无效(早期终止)的红细胞生成,这些疾病中集落形成单位-红细胞/原红细胞中的血红素过多,解释为什么这些贫血是巨红细胞性的,并表明为什么 GATA1 突变的儿童具有 DBA 样临床表型。