Department of Genetics, Institute of Gene Biology, Russian Academy of Sciences, Moscow, Russia.
Department of Molecular Biology, Princeton University, Princeton, NJ, USA.
Epigenetics Chromatin. 2019 Jan 3;12(1):2. doi: 10.1186/s13072-018-0249-2.
Boundaries in the Drosophila bithorax complex delimit autonomous regulatory domains that activate the parasegment (PS)-specific expression of homeotic genes. The Fab-7 boundary separates the iab-6 and iab-7 regulatory domains that control Abd-B expression in PS11 and PS12. This boundary is composed of multiple functionally redundant elements and has two key activities: it blocks crosstalk between iab-6 and iab-7 and facilitates boundary bypass.
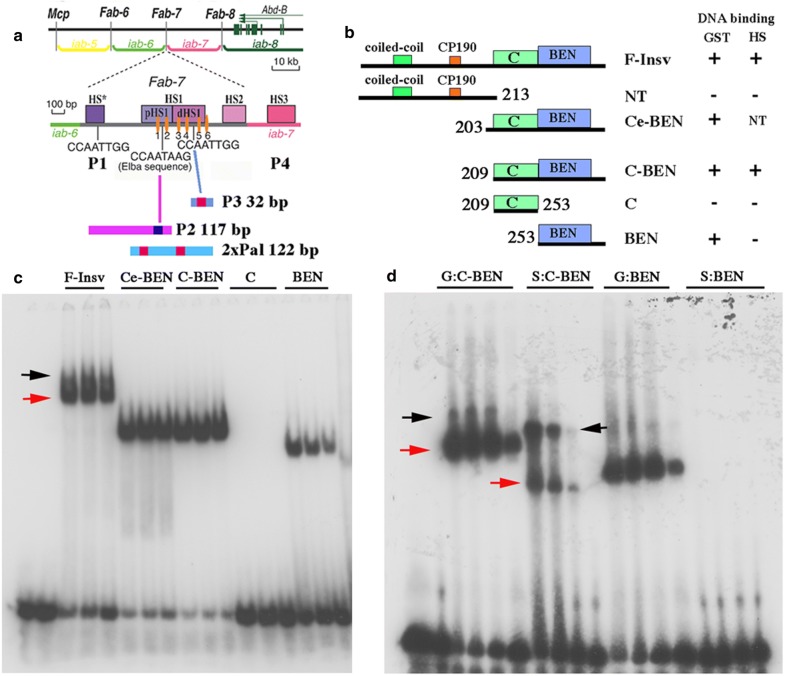
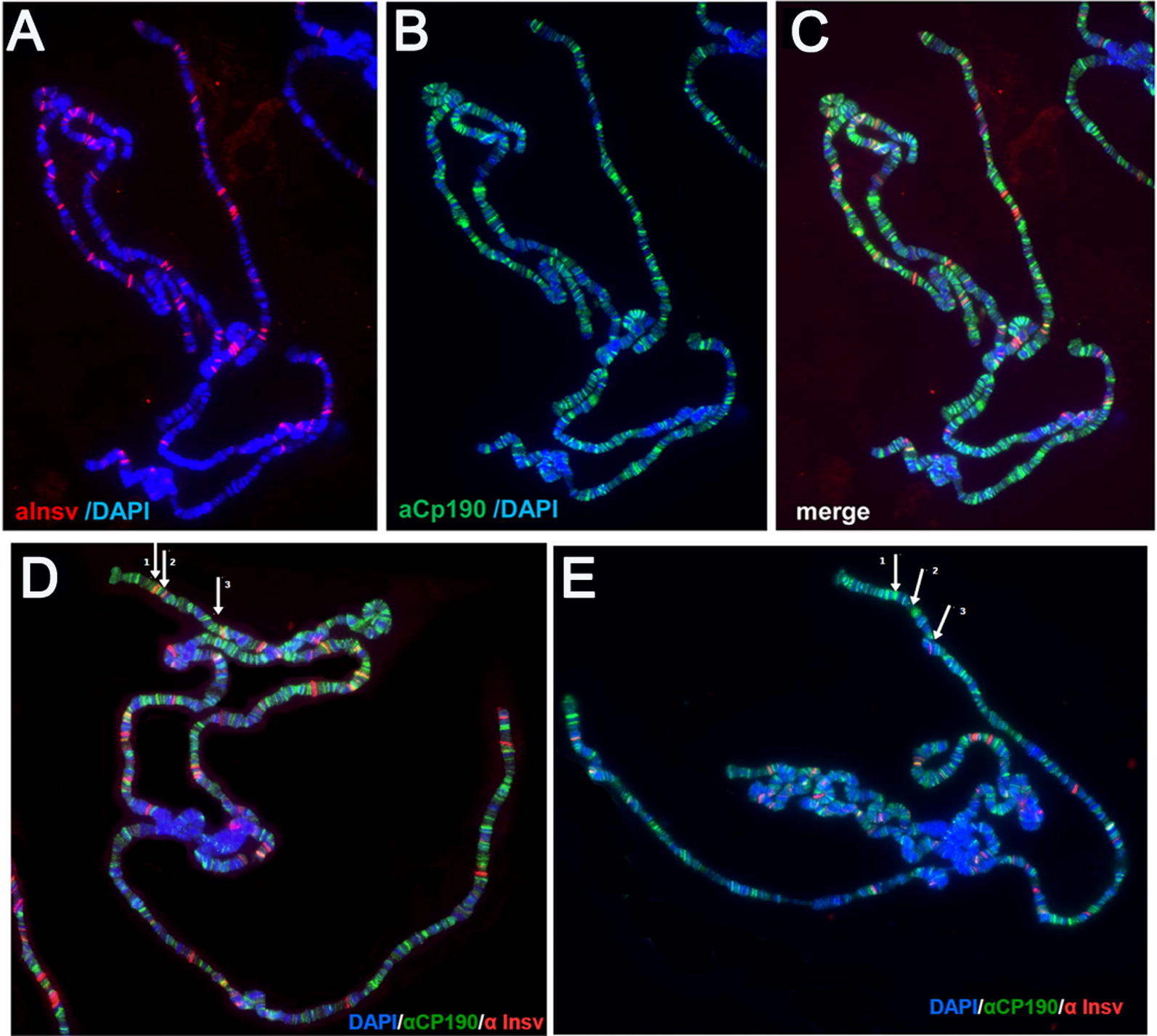
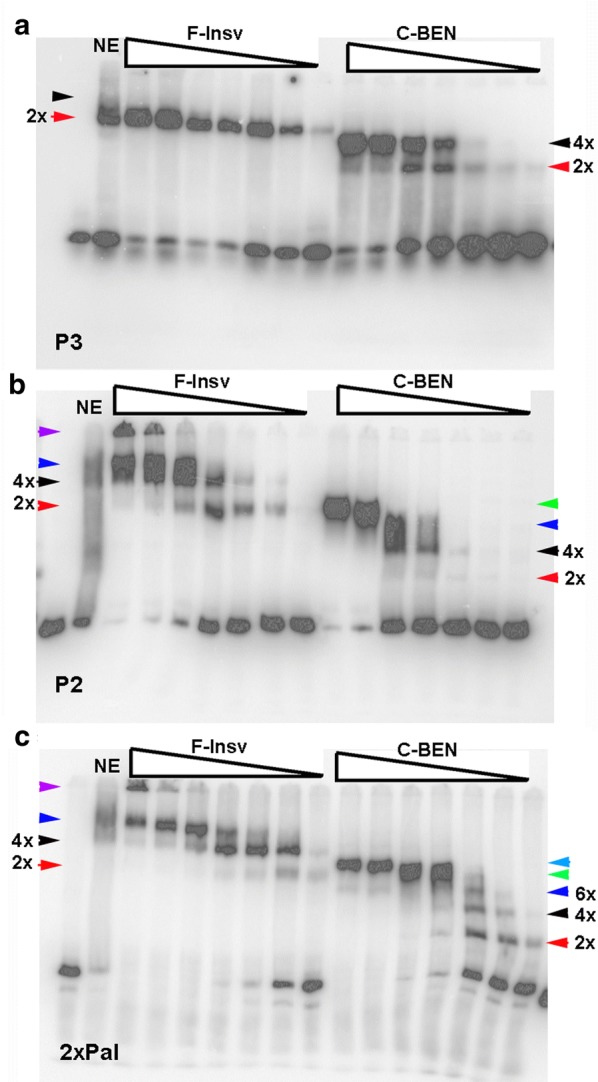
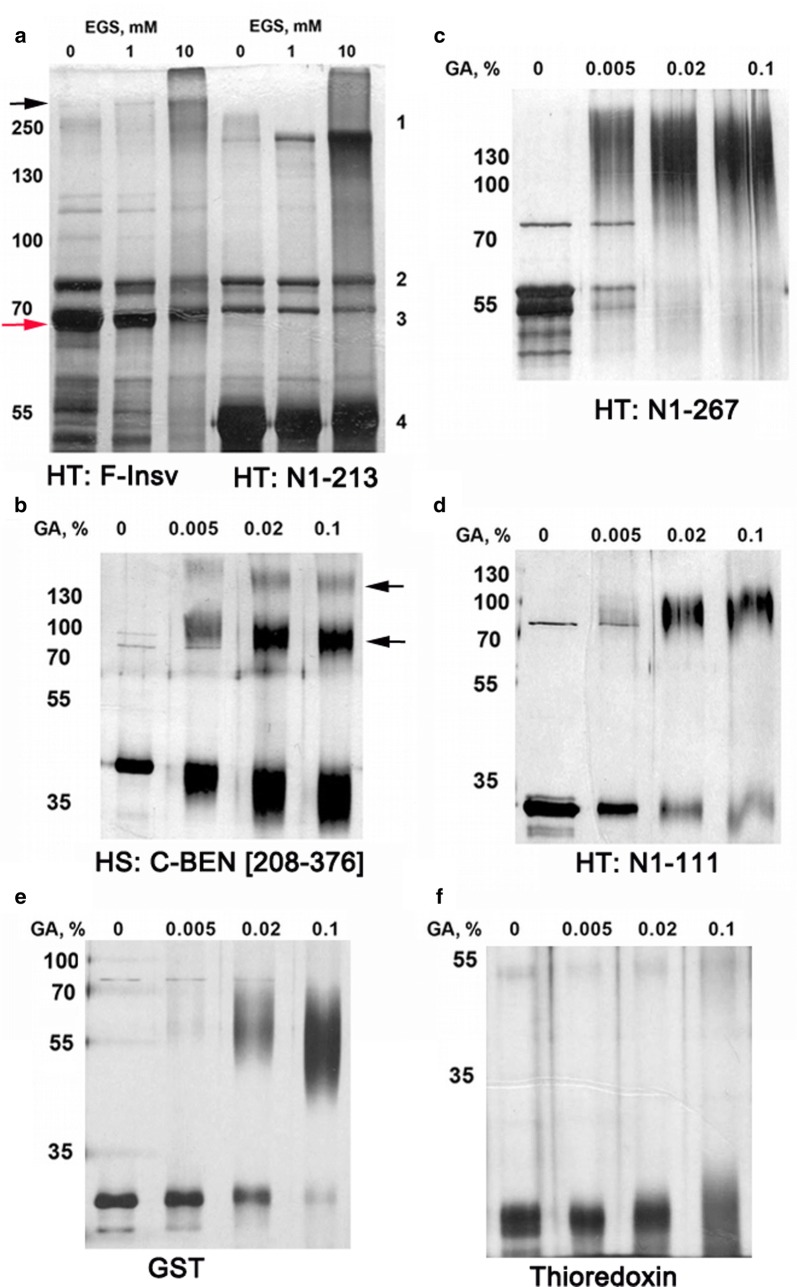
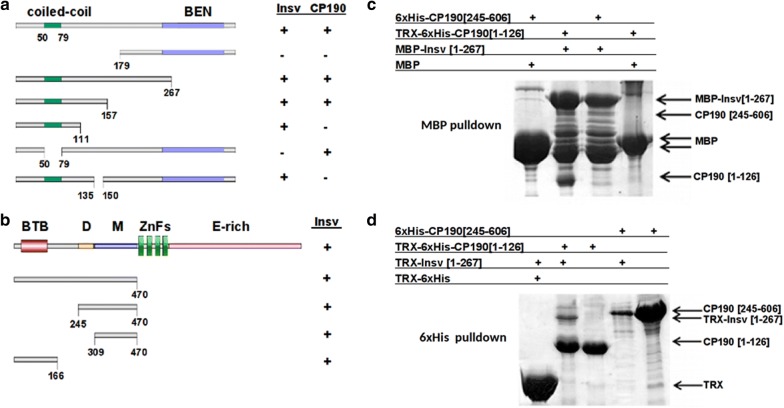
Here, we have used a structure-function approach to elucidate the biochemical properties and the in vivo activities of a conserved BEN domain protein, Insensitive, that is associated with Fab-7. Our biochemical studies indicate that in addition to the C-terminal BEN DNA-binding domain, Insv has two domains that mediate multimerization: one is a coiled-coil domain in the N-terminus, and the other is next to the BEN domain. These multimerization domains enable Insv to bind simultaneously to two canonical 8-bp recognition motifs, as well as to a ~ 100-bp non-canonical recognition sequence. They also mediate the assembly of higher-order multimers in the presence of DNA. Transgenic proteins lacking the N-terminal coiled-coil domain are compromised for boundary function in vivo. We also show that Insv interacts directly with CP190, a protein previously implicated in the boundary functions of several DNA-binding proteins, including Su(Hw) and dCTCF. While CP190 interaction is required for Insv binding to a subset of sites on polytene chromosomes, it has only a minor role in the boundary activity of Insv in the context of Fab-7.
The subdivision of eukaryotic chromosomes into discrete topological domains depends upon the pairing of boundary elements. In flies, pairing interactions are specific and typically orientation dependent. They occur in cis between neighboring heterologous boundaries, and in trans between homologous boundaries. One potential mechanism for ensuring pairing-interaction specificity is the use of sequence-specific DNA-binding proteins that can bind simultaneously with two or more recognition sequences. Our studies indicate that Insv can assemble into a multivalent DNA-binding complex and that the N-terminal Insv multimerization domain is critical for boundary function.
在果蝇的双胸复合体中,边界限定了自主调控域,这些调控域激活了同源异型基因的体节特异性表达。Fab-7 边界将 iab-6 和 iab-7 调控域分开,控制 Abd-B 在 PS11 和 PS12 中的表达。这个边界由多个功能冗余的元件组成,具有两个关键活性:它阻止了 iab-6 和 iab-7 之间的串扰,并促进了边界绕过。
在这里,我们使用结构功能方法阐明了与 Fab-7 相关的保守 BEN 结构域蛋白 Insensitive 的生化特性和体内活性。我们的生化研究表明,除了 C 末端 BEN DNA 结合结构域外,Insv 还有两个介导多聚化的结构域:一个位于 N 端的卷曲螺旋结构域,另一个位于 BEN 结构域旁边。这些多聚化结构域使 Insv 能够同时结合两个典型的 8 个碱基对识别基序,以及一个约 100 个碱基对的非典型识别序列。它们还介导了 DNA 存在时更高阶多聚体的组装。缺乏 N 端卷曲螺旋结构域的转基因蛋白在体内的边界功能受损。我们还表明,Insv 与 CP190 直接相互作用,CP190 以前被牵连到包括 Su(Hw)和 dCTCF 在内的几种 DNA 结合蛋白的边界功能中。虽然 CP190 相互作用是 Insv 结合多线染色体上某些位点所必需的,但它在 Fab-7 背景下 Insv 的边界活性中只起次要作用。
真核染色体被划分为离散的拓扑结构域,这取决于边界元件的配对。在果蝇中,配对相互作用是特异性的,通常依赖于取向。它们在邻近的异源边界之间发生顺式相互作用,在同源边界之间发生反式相互作用。确保配对相互作用特异性的一个潜在机制是使用能够同时与两个或更多识别序列结合的序列特异性 DNA 结合蛋白。我们的研究表明,Insv 可以组装成多价 DNA 结合复合物,并且 N 端 Insv 多聚化结构域对于边界功能至关重要。