Lewis Sigler Institute, Princeton University, Princeton, United States.
Department of Molecular Biology, Princeton University, Princeton, United States.
Elife. 2024 Aug 7;13:RP94070. doi: 10.7554/eLife.94070.
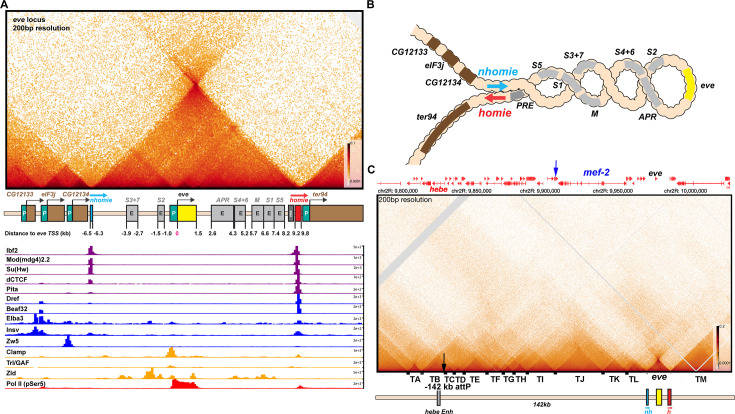
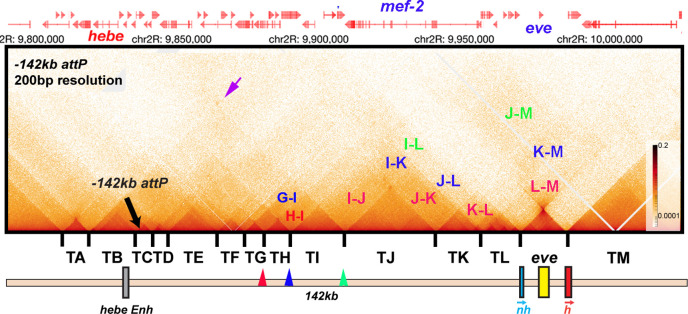
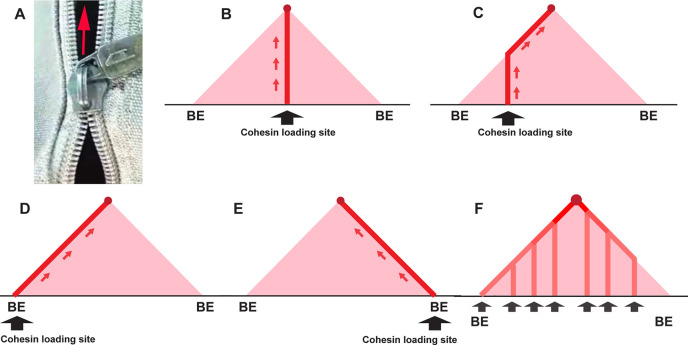
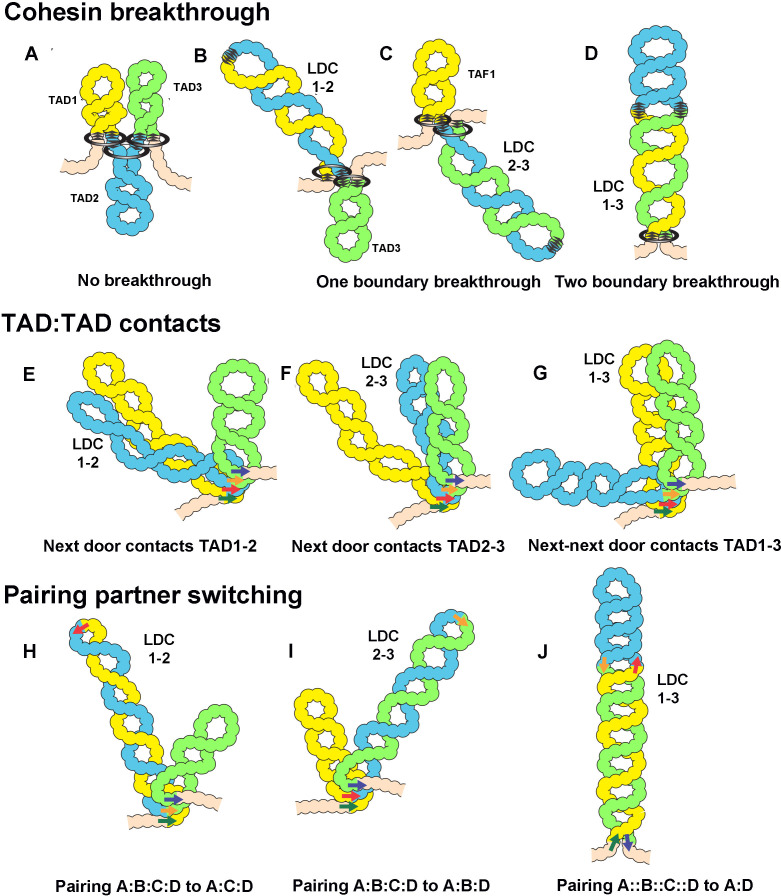
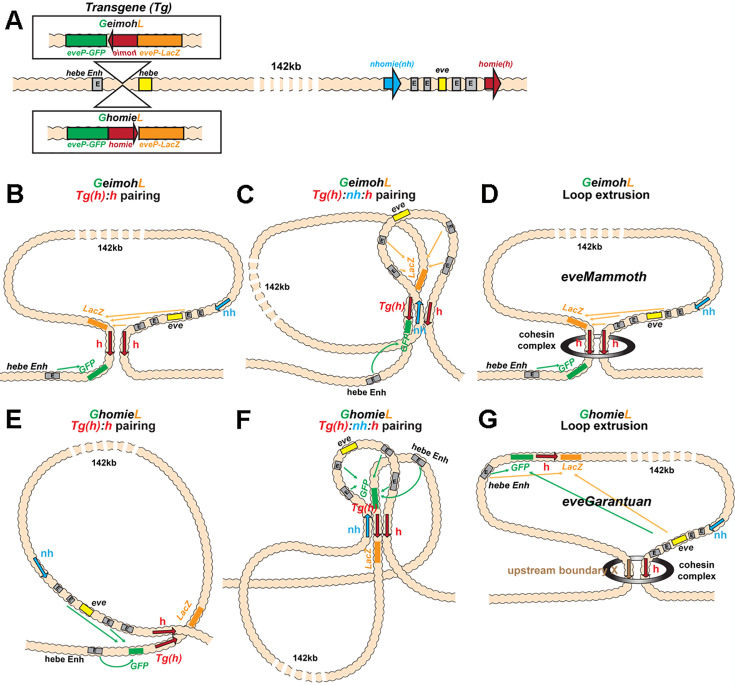
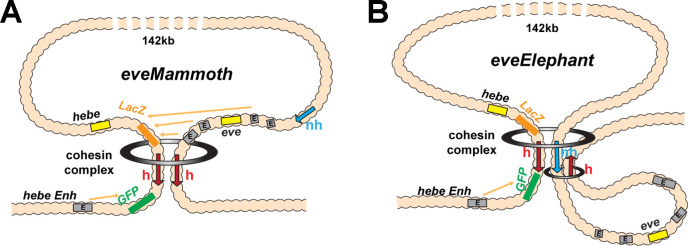
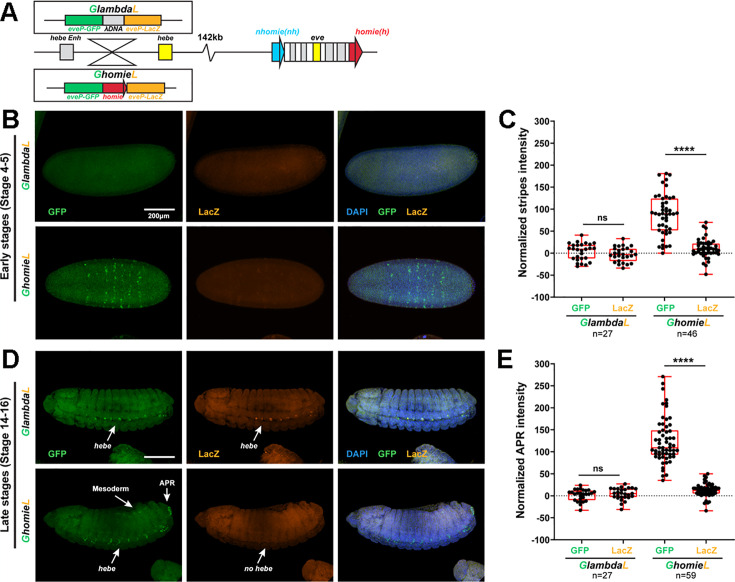
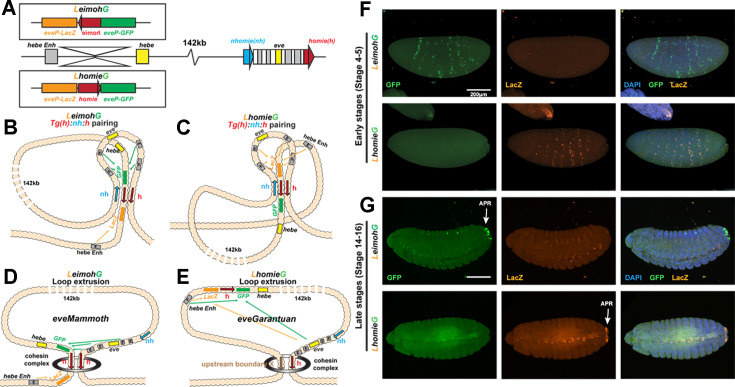
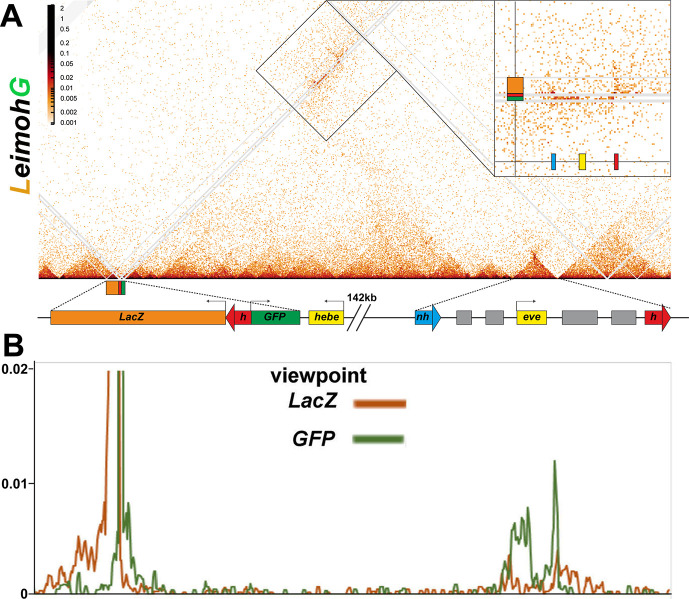
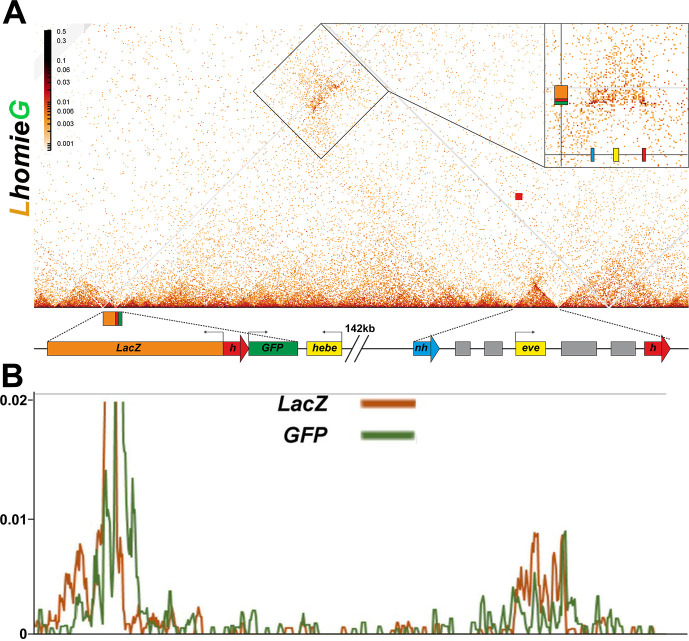
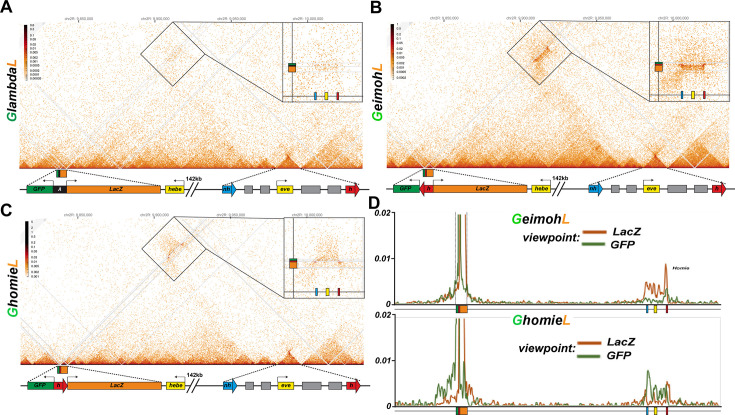
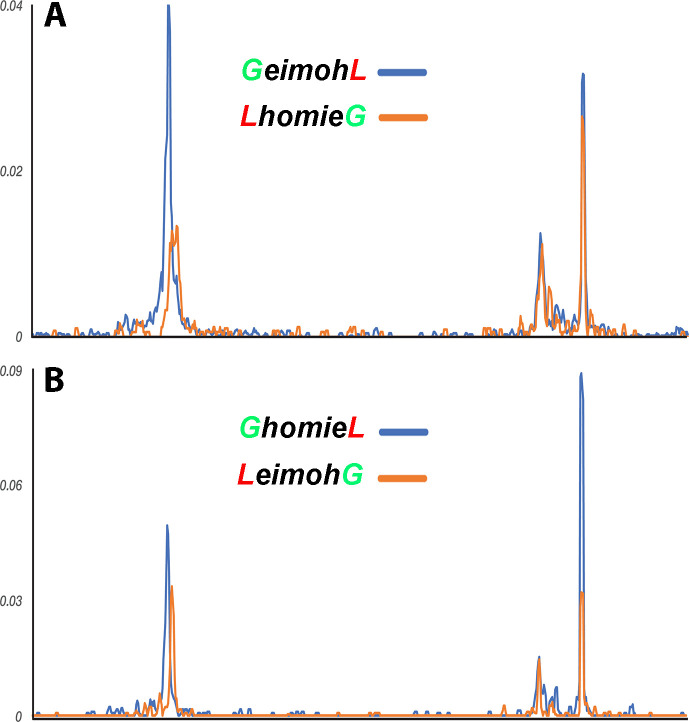
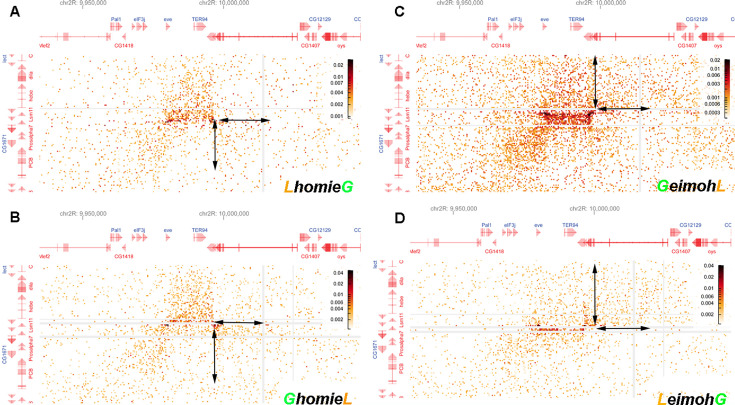
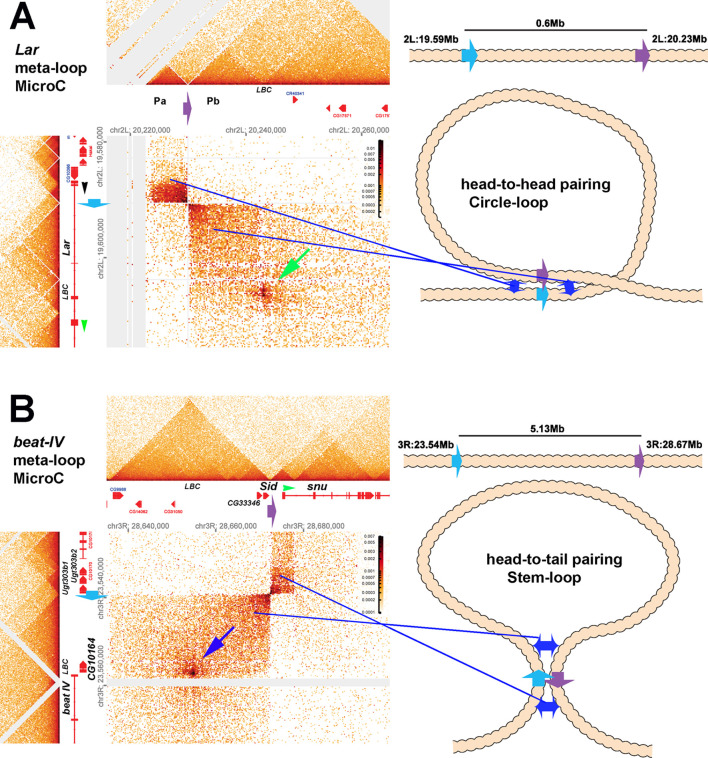
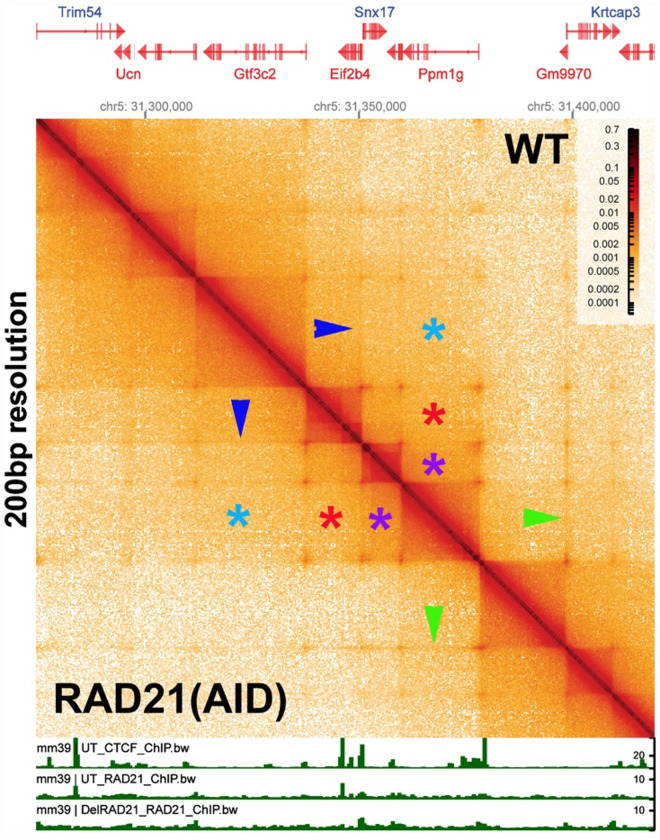
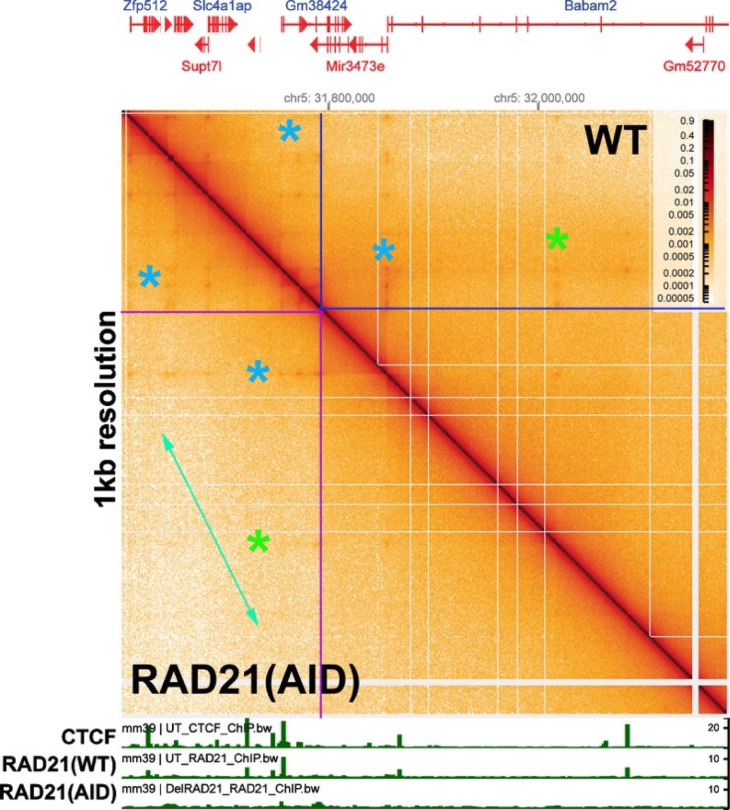
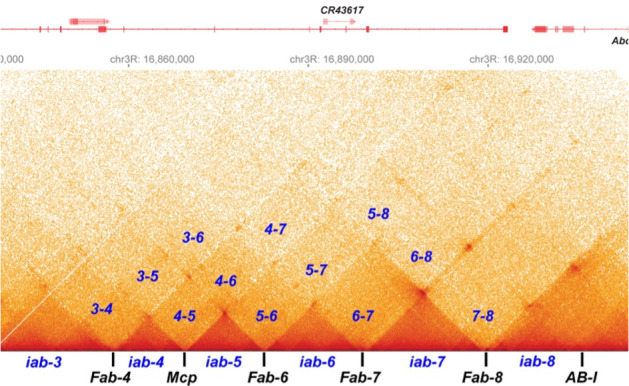
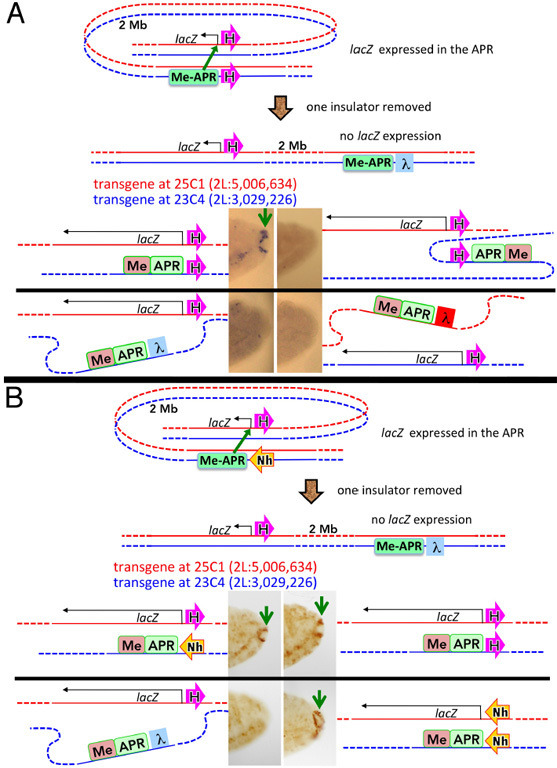
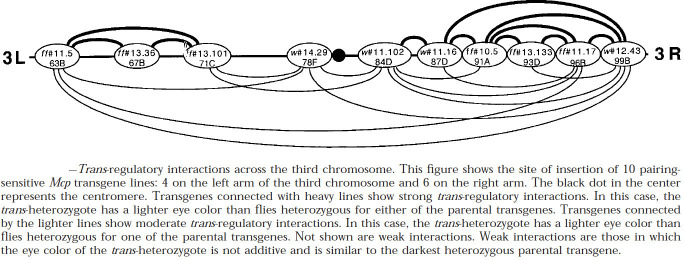
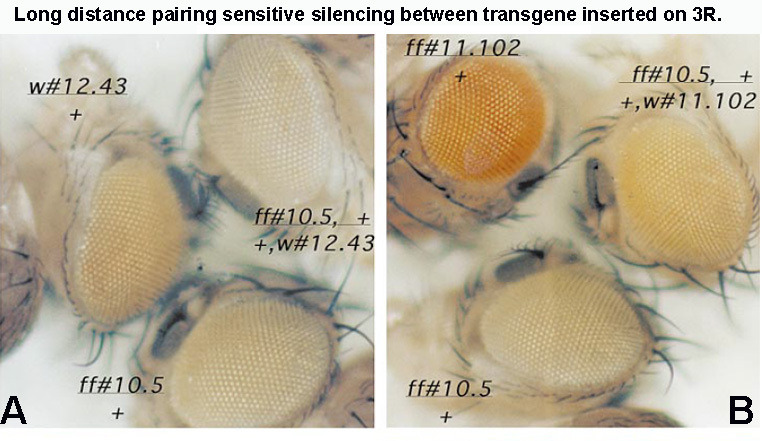
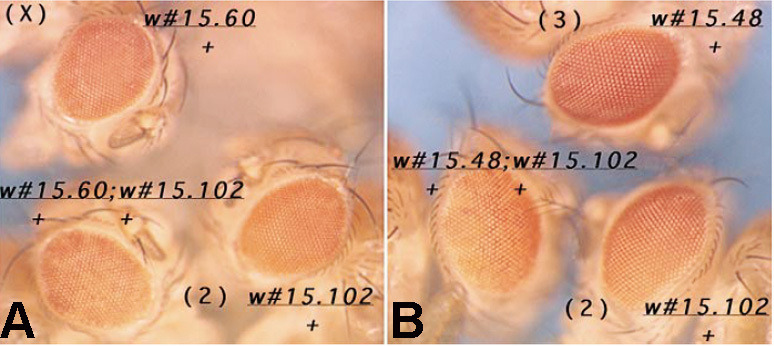
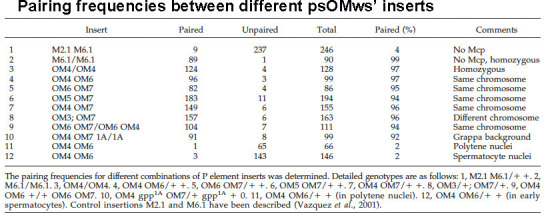
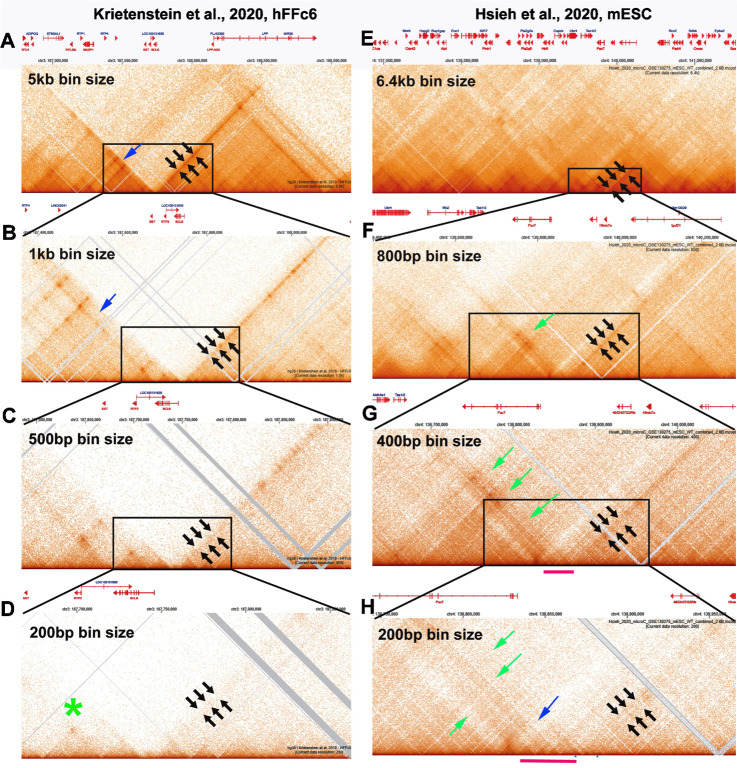
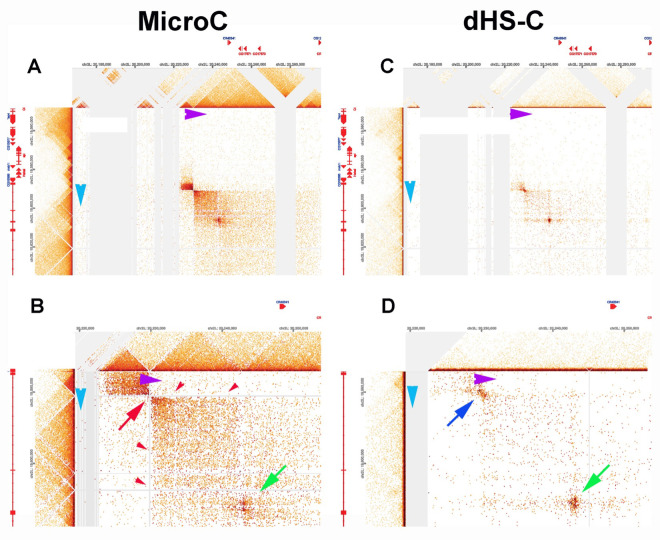
Two different models have been proposed to explain how the endpoints of chromatin looped domains ('TADs') in eukaryotic chromosomes are determined. In the first, a cohesin complex extrudes a loop until it encounters a boundary element roadblock, generating a stem-loop. In this model, boundaries are functionally autonomous: they have an intrinsic ability to halt the movement of incoming cohesin complexes that is independent of the properties of neighboring boundaries. In the second, loops are generated by boundary:boundary pairing. In this model, boundaries are functionally non-autonomous, and their ability to form a loop depends upon how well they match with their neighbors. Moreover, unlike the loop-extrusion model, pairing interactions can generate both stem-loops and circle-loops. We have used a combination of MicroC to analyze how TADs are organized, and experimental manipulations of the TAD boundary, , to test the predictions of the 'loop-extrusion' and the 'boundary-pairing' models. Our findings are incompatible with the loop-extrusion model, and instead suggest that the endpoints of TADs in flies are determined by a mechanism in which boundary elements physically pair with their partners, either head-to-head or head-to-tail, with varying degrees of specificity. Although our experiments do not address how partners find each other, the mechanism is unlikely to require loop extrusion.
两种不同的模型被提出来解释真核染色体染色质环域('TADs')的末端是如何确定的。在第一种模型中,一个黏合复合物伸出一个环,直到遇到一个边界元件障碍,产生一个茎环。在这个模型中,边界是功能自主的:它们具有内在的能力来阻止进入的黏合复合物的运动,这种能力独立于相邻边界的性质。在第二种模型中,环是由边界:边界配对产生的。在这个模型中,边界是功能非自主的,它们形成环的能力取决于它们与邻居的匹配程度。此外,与环伸出模型不同,配对相互作用可以产生茎环和圆形环。我们使用 MicroC 的组合来分析 TADs 是如何组织的,以及对 TAD 边界的实验操作,以检验 '环伸出'和 '边界配对'模型的预测。我们的发现与环伸出模型不一致,而是表明果蝇 TADs 的末端是由一种机制决定的,在这种机制中,边界元件与它们的伙伴物理配对,要么头对头,要么头对尾,具有不同程度的特异性。尽管我们的实验没有解决伙伴如何找到彼此的问题,但该机制不太可能需要环伸出。