Johnson Kenneth A
Department of Molecular Biosciences, The University of Texas, Austin, TX 78735, USA.
Beilstein J Org Chem. 2019 Jan 2;15:16-29. doi: 10.3762/bjoc.15.2. eCollection 2019.
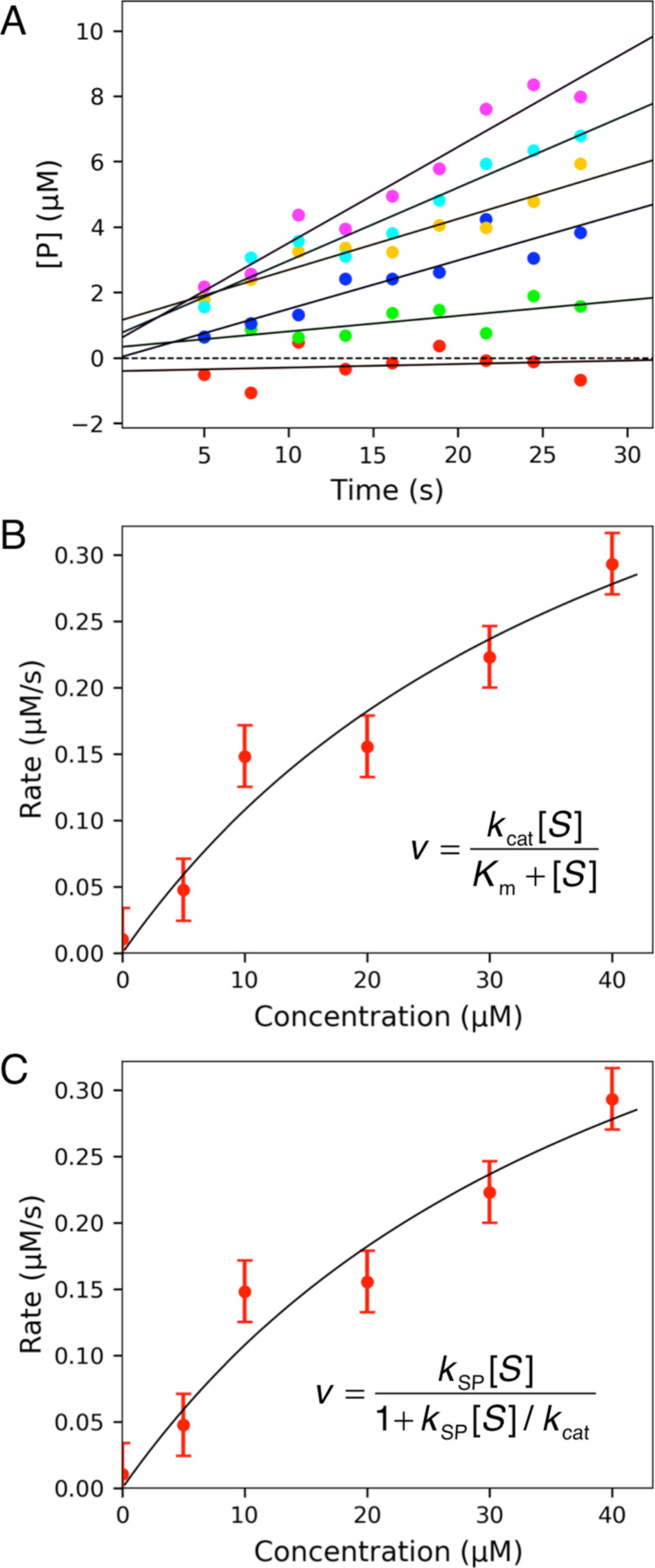
The Michaelis-Menten equation is usually expressed in terms of and values: = [S]/( + [S]). However, it is impossible to interpret in the absence of additional information, while the ratio of / provides a measure of enzyme specificity and is proportional to enzyme efficiency and proficiency. Moreover, / provides a lower limit on the second order rate constant for substrate binding. For these reasons it is better to redefine the Michaelis-Menten equation in terms of and / values: = [S]/(1 + [S]/ ), where the specificity constant, = / . In this short review, the rationale for this assertion is explained and it is shown that more accurate measurements of / can be derived directly using the modified form of the Michaelis-Menten equation rather than calculated from the ratio of and values measured separately. Even greater accuracy is achieved with fitting the raw data directly by numerical integration of the rate equations rather than using analytically derived equations. The importance of fitting to derive and / is illustrated by considering the role of conformational changes in enzyme specificity where and / can reflect different steps in the pathway. This highlights the pitfalls in attempting to interpret , which is best understood as the ratio of divided by / .
米氏方程通常用(K_m)和(V_{max})值表示:(V = V_{max}[S]/(K_m + [S]))。然而,在没有额外信息的情况下,无法解释(K_m),而(V_{max}/K_m)提供了一种衡量酶特异性的指标,并且与酶的效率和效能成正比。此外,(V_{max}/K_m)为底物结合的二级速率常数提供了一个下限。基于这些原因,最好根据(K_m)和(V_{max}/K_m)值重新定义米氏方程:(V = V_{max}[S]/(1 + K_m [S]/V_{max})),其中特异性常数(k_{cat}/K_m = V_{max}/K_m)。在这篇简短的综述中,解释了这一断言的基本原理,并表明可以直接使用米氏方程的修正形式更准确地测量(V_{max}/K_m),而不是从分别测量的(V_{max})和(K_m)值的比值计算得出。通过直接对速率方程进行数值积分拟合原始数据,而不是使用解析推导的方程,可以获得更高的准确性。通过考虑构象变化在酶特异性中的作用来说明拟合以得出(K_m)和(V_{max}/K_m)的重要性,其中(K_m)和(V_{max}/K_m)可以反映途径中的不同步骤。这突出了试图解释(K_m)时的陷阱,(K_m)最好理解为(V_{max})除以(V_{max}/K_m)的比值。