Opperman Diederik Johannes, Murgida Daniel Horacio, Dalosto Sergio Daniel, Brondino Carlos Dante, Ferroni Felix Martín
Department of Biotechnology, University of the Free State, 205 Nelson Mandela Drive, Bloemfontein, Free State 9300, South Africa.
Departamento de Química Inorgánica, Analítica y Química Física and INQUIMAE (CONICET-UBA), Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria, Pab. 2 piso 1, Buenos Aires, Buenos Aires C1428EHA, Argentina.
IUCrJ. 2019 Feb 9;6(Pt 2):248-258. doi: 10.1107/S2052252519000241. eCollection 2019 Mar 1.
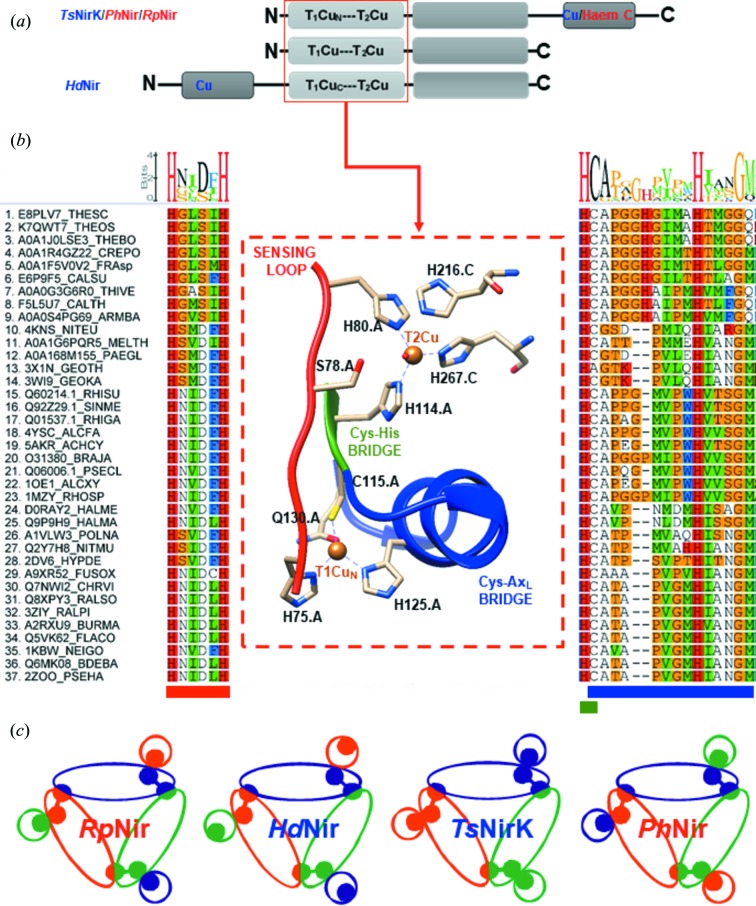
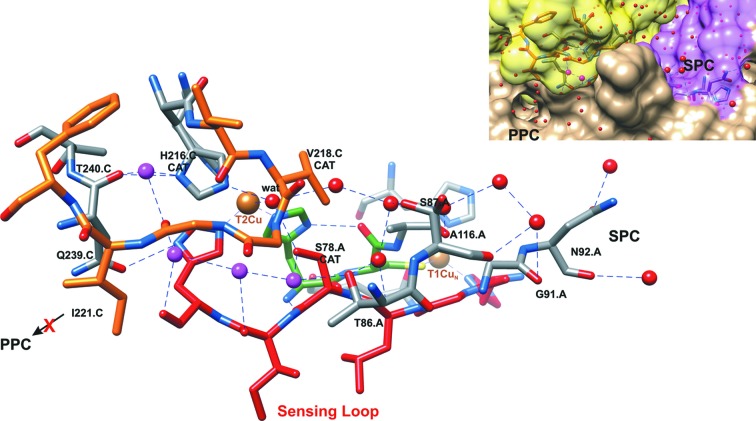
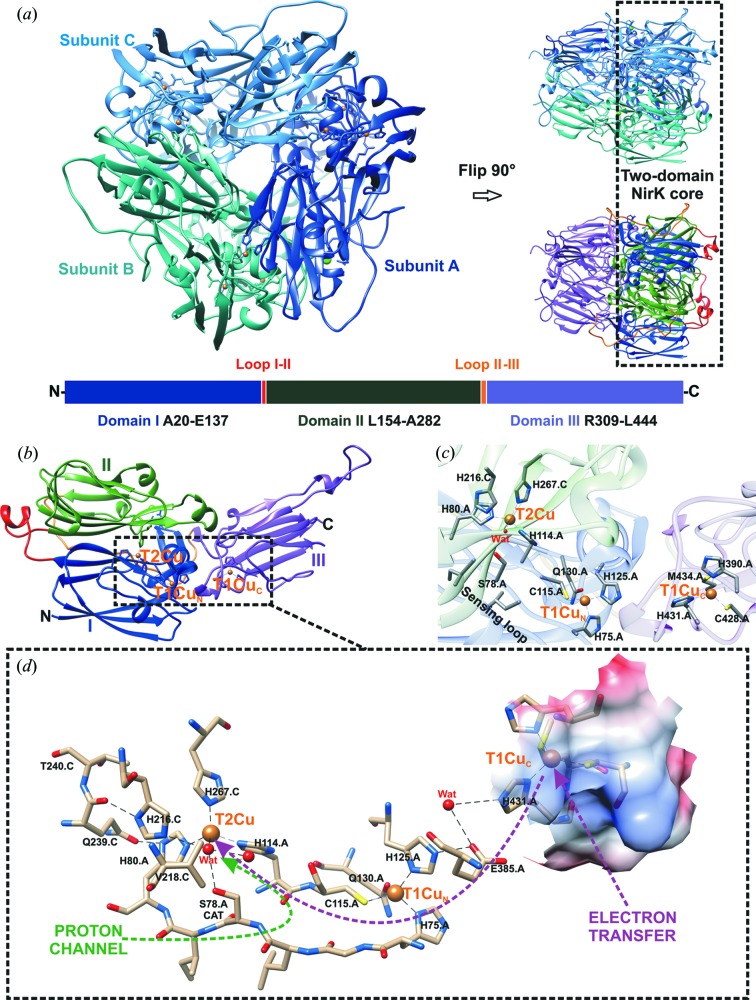
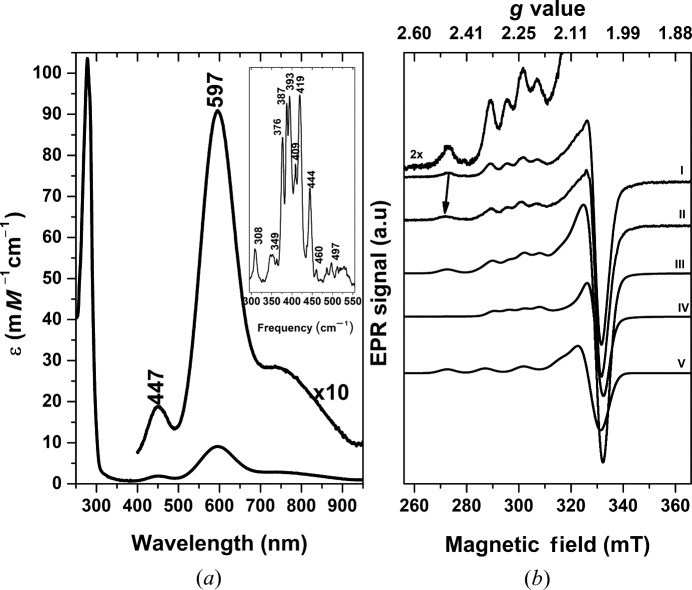
Dissimilatory nitrite reductases are key enzymes in the denitrification pathway, reducing nitrite and leading to the production of gaseous products (NO, NO and N). The reaction is catalysed either by a Cu-containing nitrite reductase (NirK) or by a cytochrome nitrite reductase (NirS), as the simultaneous presence of the two enzymes has never been detected in the same microorganism. The thermophilic bacterium SA-01 is an exception to this rule, harbouring both genes within a denitrification cluster, which encodes for an atypical NirK. The crystal structure of NirK has been determined at 1.63 Å resolution. NirK is a homotrimer with subunits of 451 residues that contain three copper atoms each. The N-terminal region possesses a type 2 Cu (T2Cu) and a type 1 Cu (T1Cu) while the C-terminus contains an extra type 1 Cu (T1Cu) bound within a cupredoxin motif. T1Cu shows an unusual Cu atom coordination (His-Cys-Gln) compared with T1Cu observed in NirKs reported so far (His-Cys-Met). T1Cu is buried at ∼5 Å from the molecular surface and located ∼14.1 Å away from T1Cu; T1Cu and T2Cu are ∼12.6 Å apart. All these distances are compatible with an electron-transfer process T1Cu → T1Cu → T2Cu. T1Cu and T2Cu are connected by a typical Cys-His bridge and an unexpected sensing loop which harbours a Ser residue close to T2Cu, suggesting an alternative nitrite-reduction mechanism in these enzymes. Biophysicochemical and functional features of NirK are discussed on the basis of X-ray crystallography, electron paramagnetic resonance, resonance Raman and kinetic experiments.
异化亚硝酸盐还原酶是反硝化途径中的关键酶,可还原亚硝酸盐并导致气态产物(NO、NO和N)的产生。该反应由含铜亚硝酸盐还原酶(NirK)或细胞色素亚硝酸盐还原酶(NirS)催化,因为从未在同一微生物中同时检测到这两种酶。嗜热细菌SA-01是该规则的一个例外,在一个反硝化簇中同时含有这两个基因,该簇编码一种非典型的NirK。已确定NirK的晶体结构,分辨率为1.63 Å。NirK是一种同三聚体,亚基有451个残基,每个亚基含有三个铜原子。N端区域含有一个2型铜(T2Cu)和一个1型铜(T1Cu),而C端含有一个额外的1型铜(T1Cu),结合在一个铜蓝蛋白基序内。与迄今为止报道的NirK中观察到的T1Cu(His-Cys-Met)相比,T1Cu显示出不寻常的铜原子配位(His-Cys-Gln)。T1Cu埋在距分子表面约5 Å处,距离T1Cu约14.1 Å;T1Cu和T2Cu相距约12.6 Å。所有这些距离都与电子转移过程T1Cu → T1Cu → T2Cu相符合。T1Cu和T2Cu通过一个典型的Cys-His桥和一个意想不到的传感环相连,该传感环在靠近T2Cu处有一个Ser残基,表明这些酶中存在一种替代的亚硝酸盐还原机制。基于X射线晶体学、电子顺磁共振、共振拉曼和动力学实验,讨论了NirK的生物物理化学和功能特征。