Biophysics Graduate Program, The Ohio State University, Columbus, United States.
Department of Biochemistry and Molecular Biology, The Pennsylvania State University, State College, United States.
Elife. 2019 Mar 19;8:e43008. doi: 10.7554/eLife.43008.
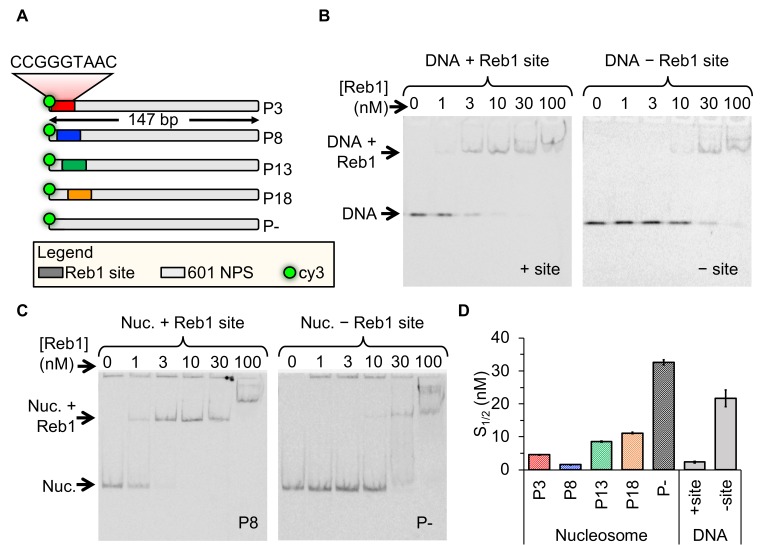
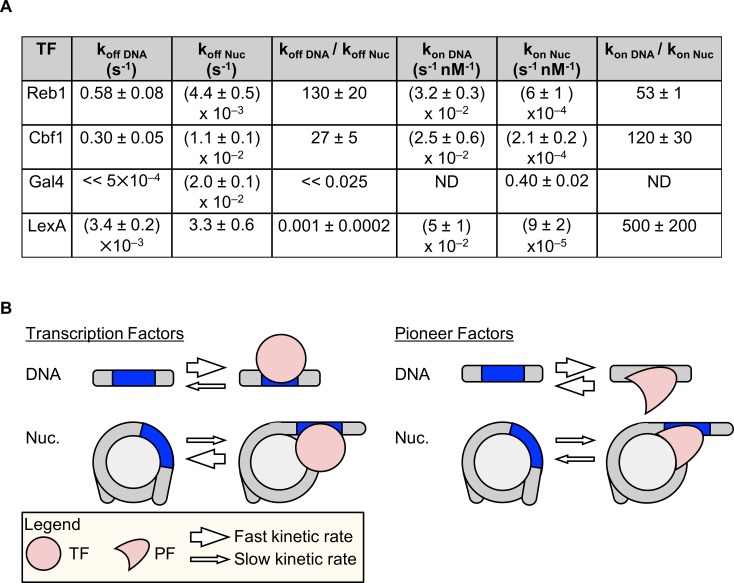
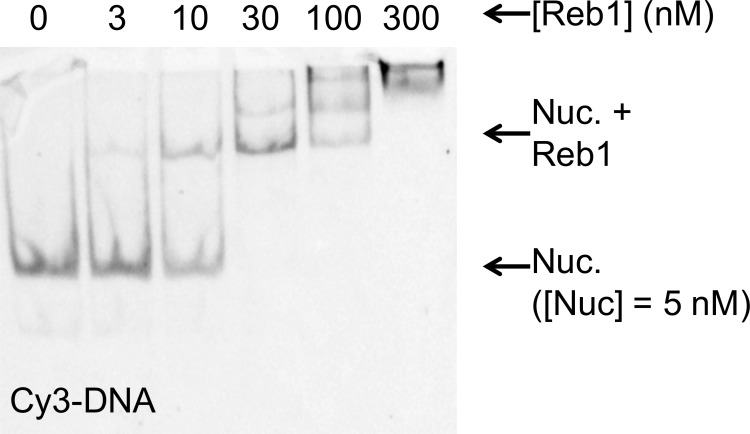
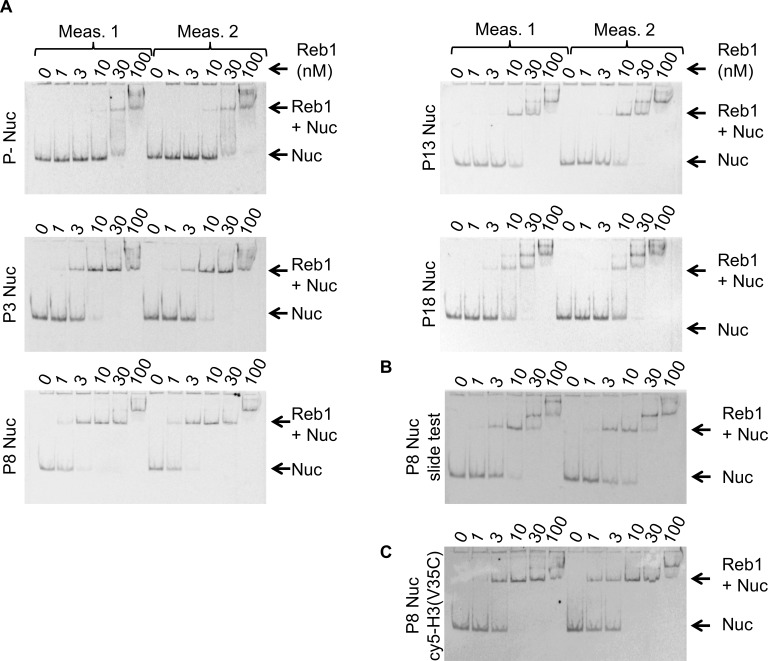
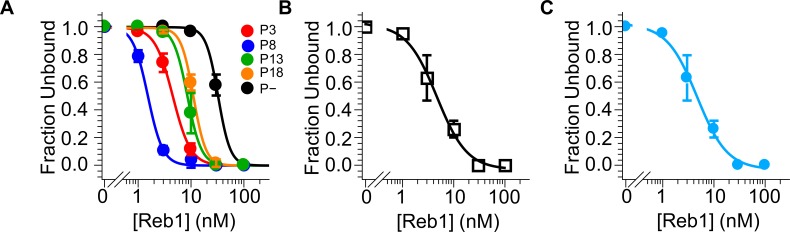
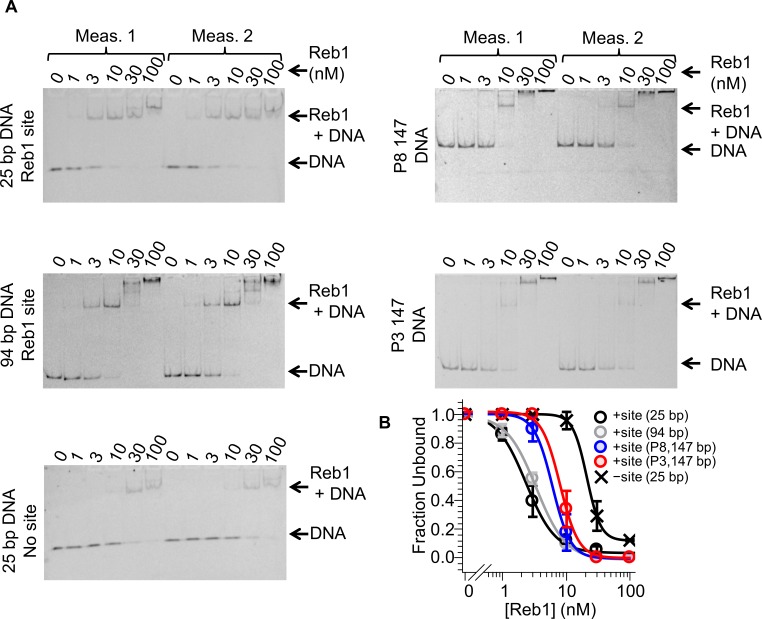
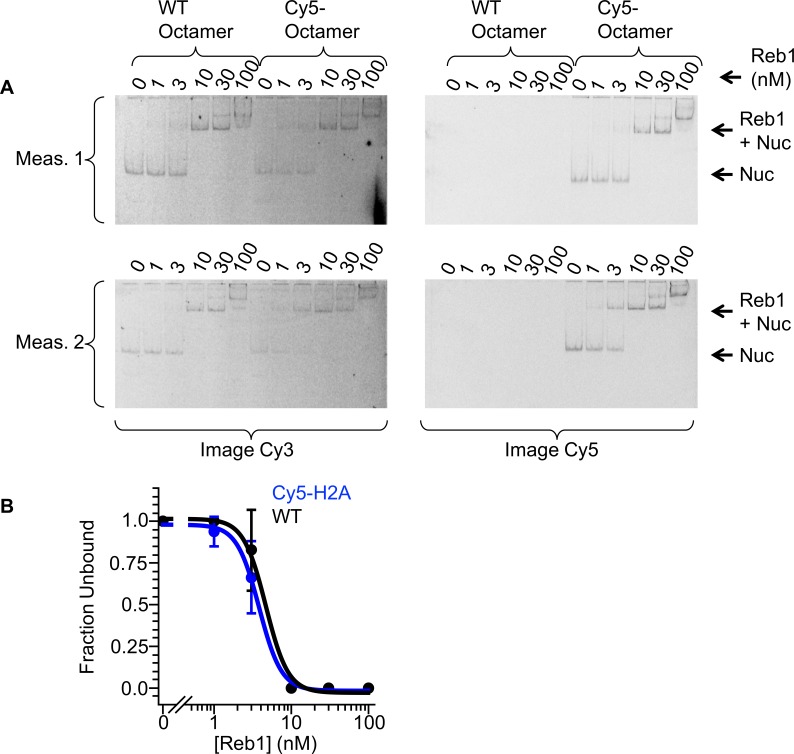
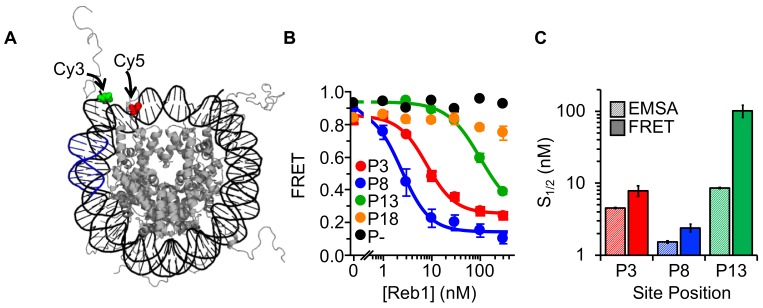
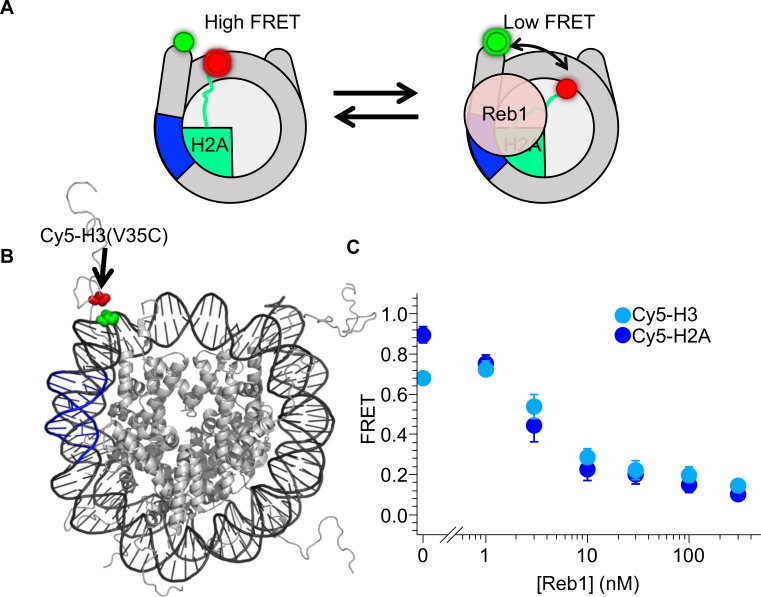
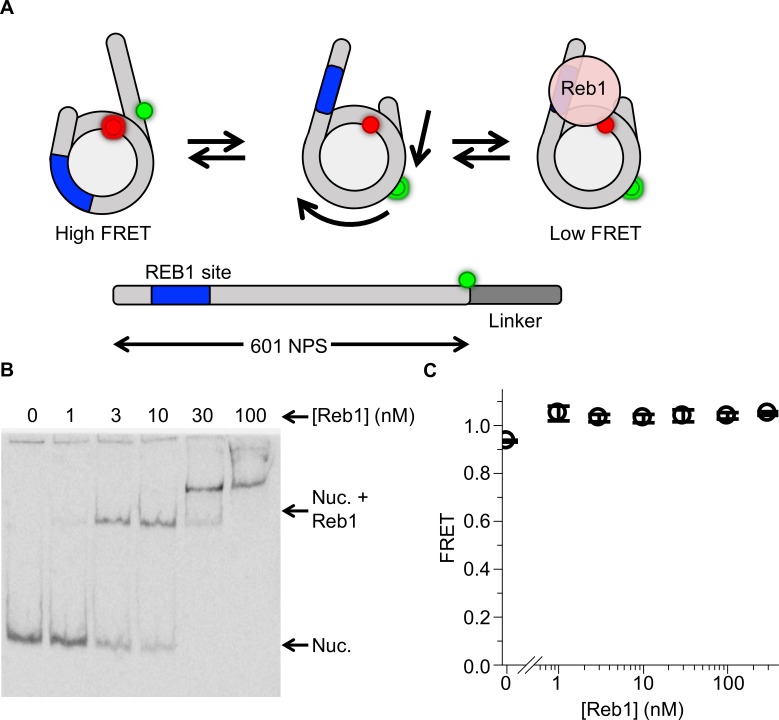
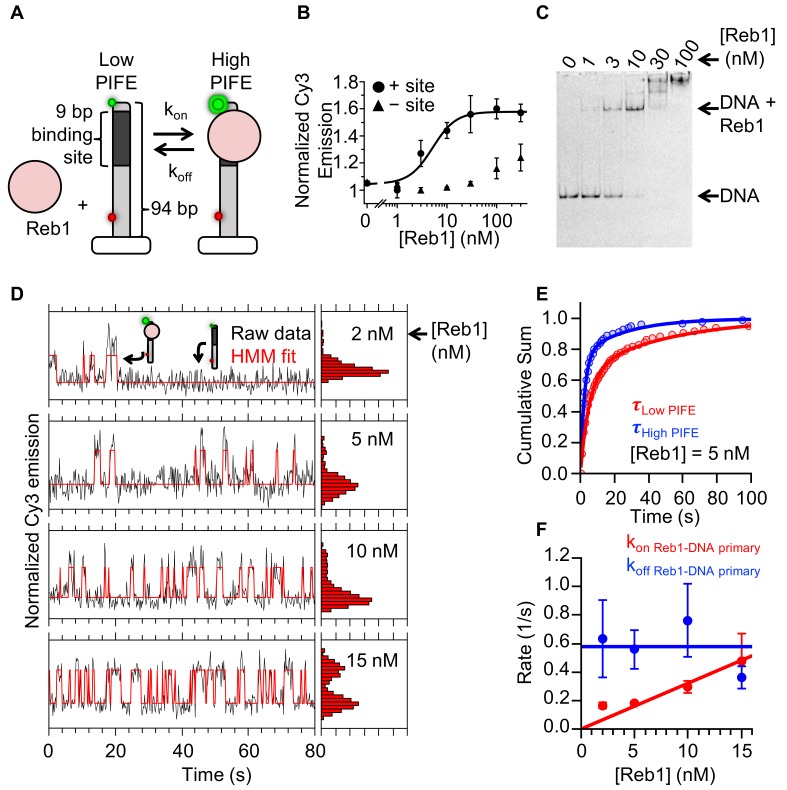
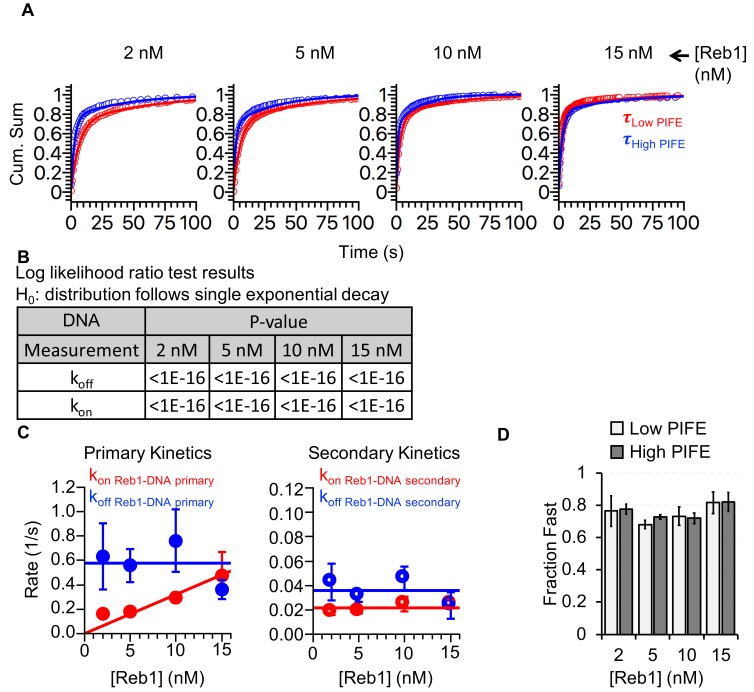
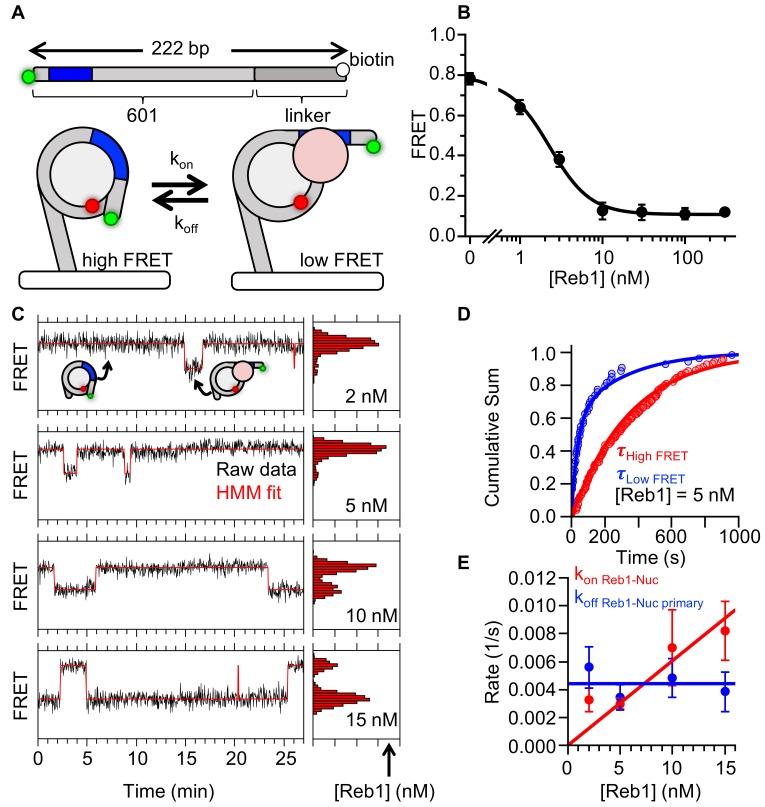
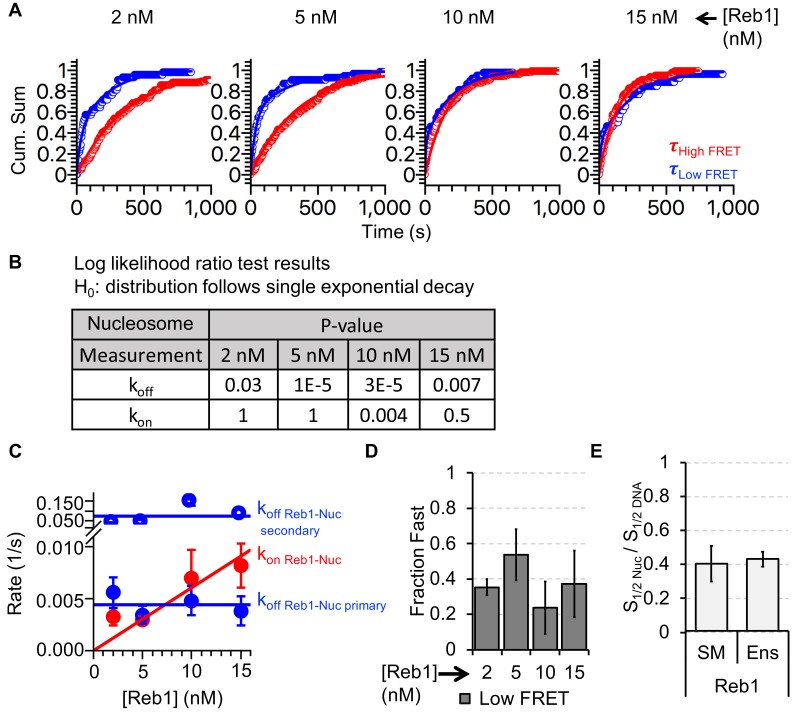
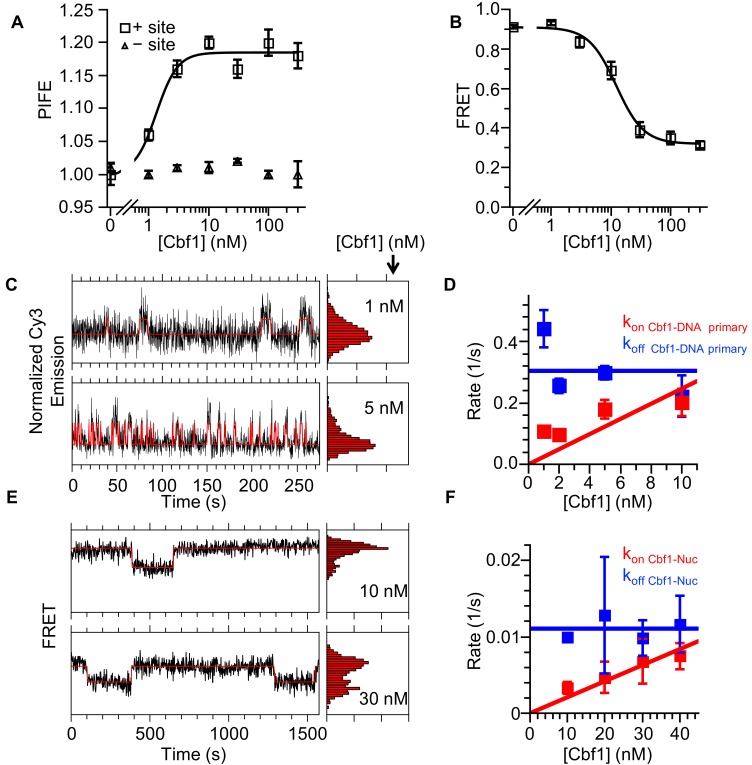
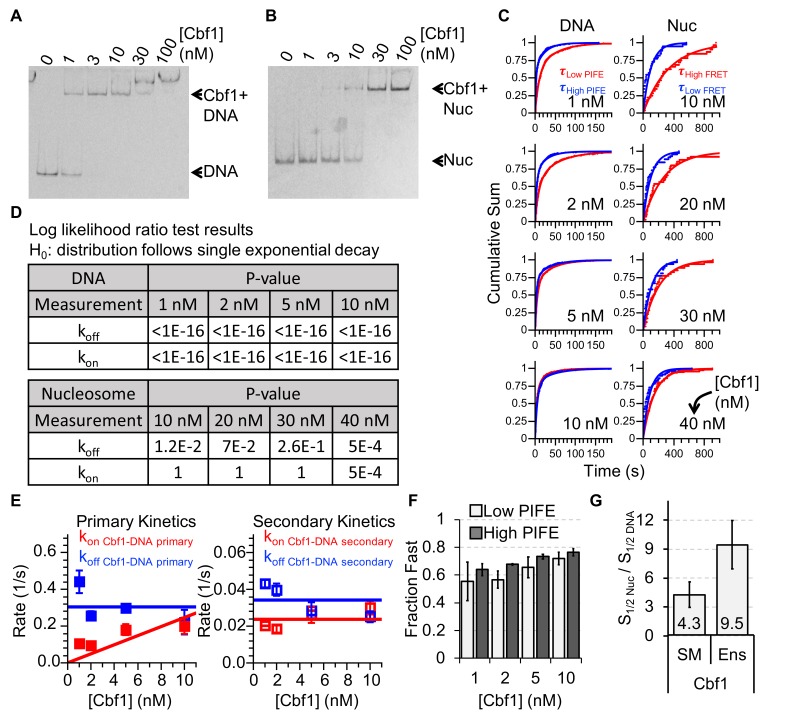
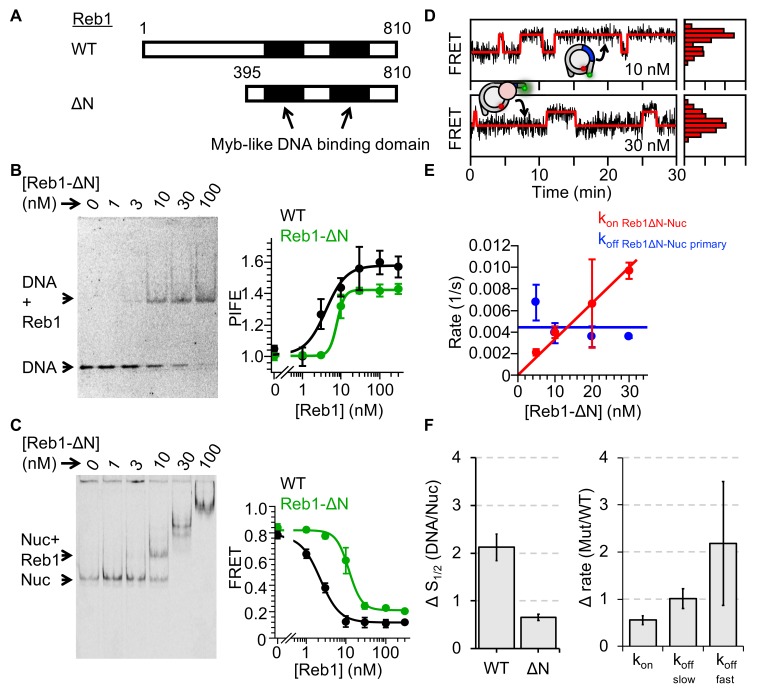
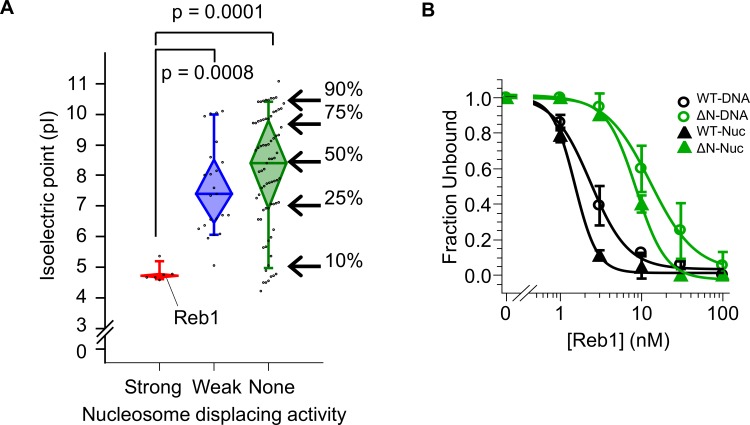
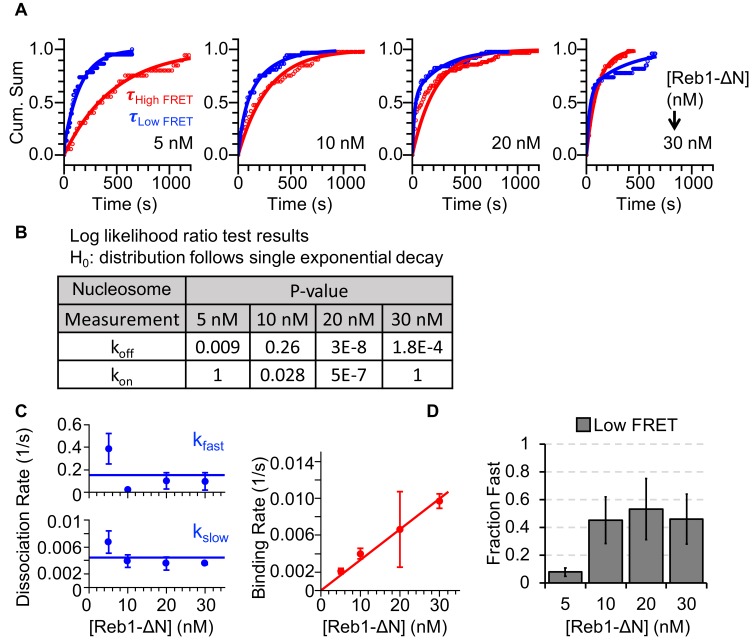
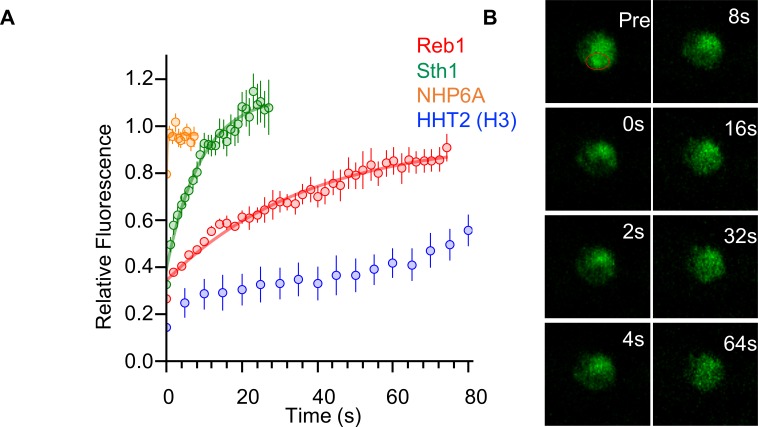
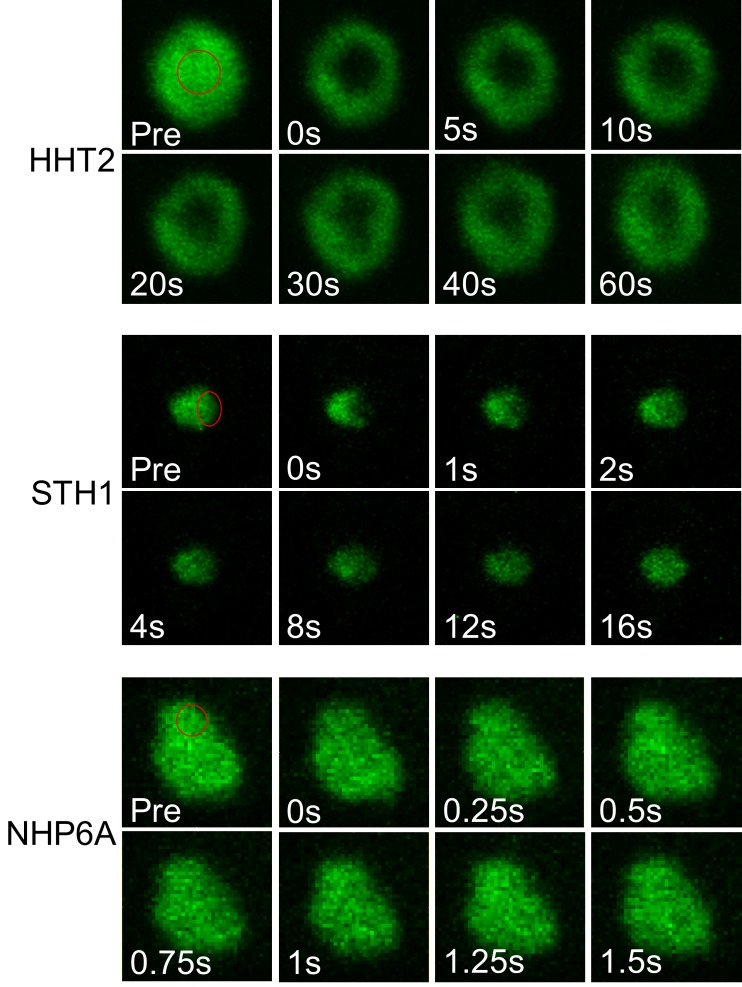
Nucleosomes restrict the occupancy of most transcription factors (TF) by reducing binding and accelerating dissociation, while a small group of TFs have high affinities to nucleosome-embedded sites and facilitate nucleosome displacement. To understand this process mechanistically, we investigated two TFs, Reb1 and Cbf1. We show that these factors bind to their sites within nucleosomes with similar binding affinities as to naked DNA, trapping a partially unwrapped nucleosome without histone eviction. Both the binding and dissociation rates of Reb1 and Cbf1 are significantly slower at the nucleosomal sites relative to those for naked DNA, demonstrating that the high affinities are achieved by increasing the dwell time on nucleosomes in order to compensate for reduced binding. Reb1 also shows slow migration rate in the yeast nuclei. These properties are similar to those of human pioneer factors (PFs), suggesting that the mechanism of nucleosome targeting is conserved from yeast to humans.
核小体通过降低结合亲和力和加速解离来限制大多数转录因子 (TF) 的占有率,而一小部分 TF 与核小体结合的亲和力很高,并能促进核小体位移。为了从机制上理解这一过程,我们研究了两个 TF,Reb1 和 Cbf1。我们发现这些因子与核小体中的结合位点的结合亲和力与裸露 DNA 相似,在没有组蛋白逐出的情况下捕获部分解缠的核小体。Reb1 和 Cbf1 的结合和解离速率在核小体上相对于裸露 DNA 显著降低,表明高亲和力是通过增加在核小体上的停留时间来实现的,以补偿结合的减少。Reb1 在酵母核中也表现出缓慢的迁移率。这些特性与人类先驱因子 (PFs) 的特性相似,这表明从酵母到人,核小体靶向的机制是保守的。