Max Planck Institute of Biochemistry, Molecular Cell Biology, Martinsried, Germany.
Biomedizinisches Centrum, Core Facility Bioinformatics, Ludwig-Maximilians-Universität München, Martinsried, Germany.
EMBO J. 2019 Jun 3;38(11). doi: 10.15252/embj.2018100368. Epub 2019 Apr 23.
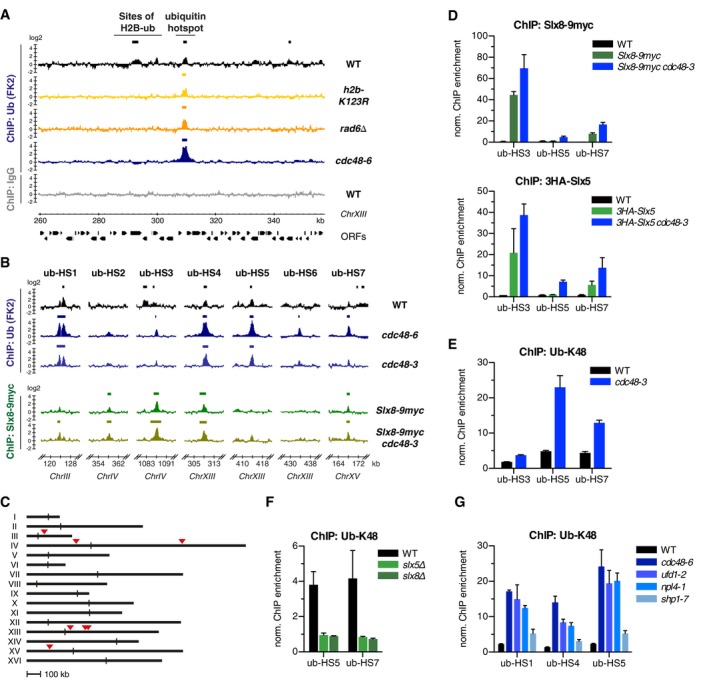
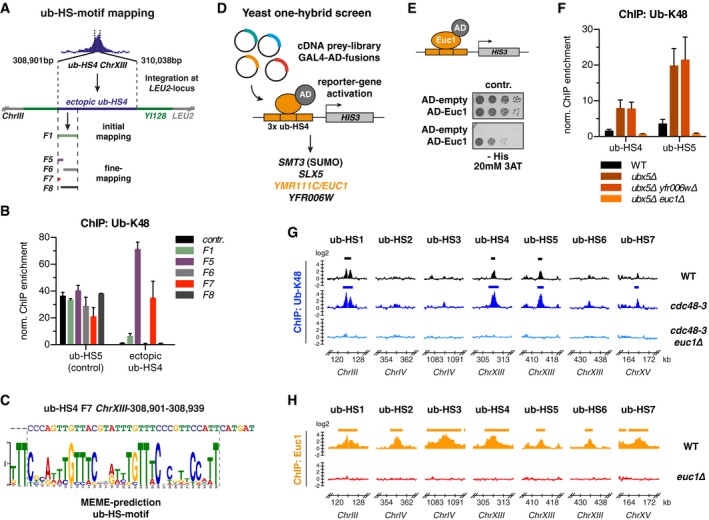
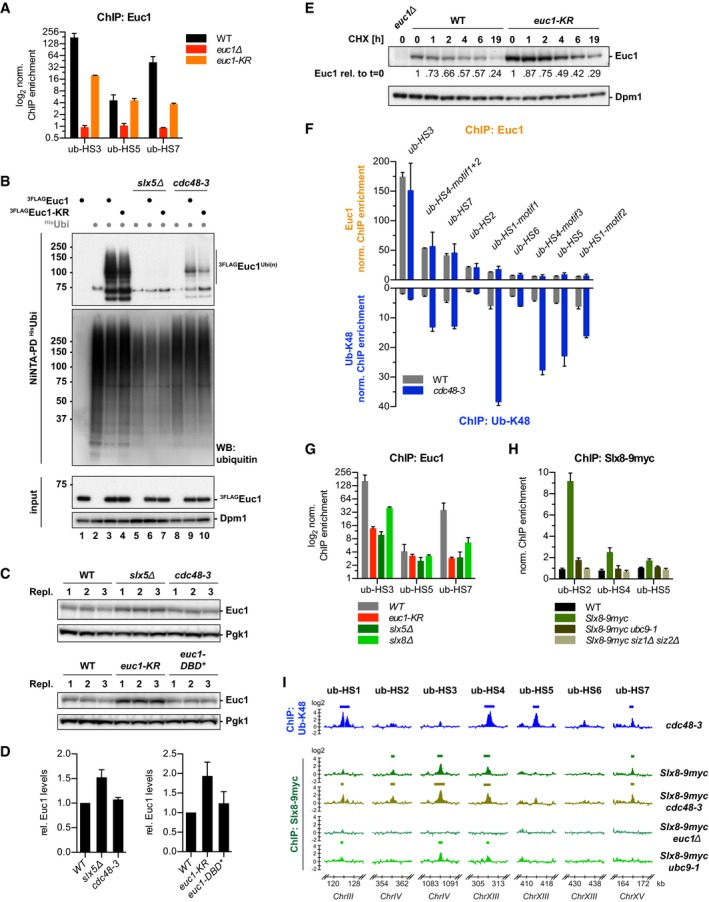
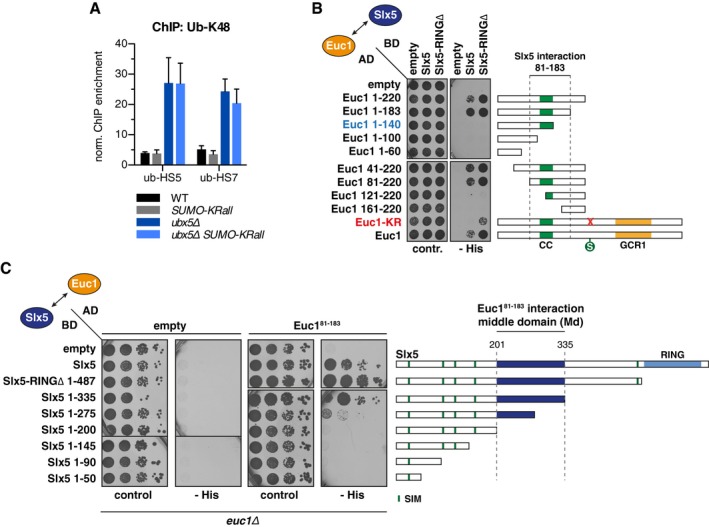
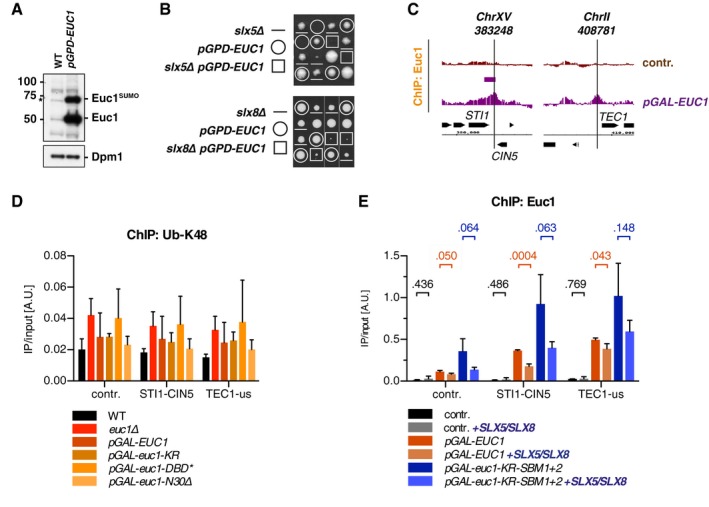
Chromatin is a highly regulated environment, and protein association with chromatin is often controlled by post-translational modifications and the corresponding enzymatic machinery. Specifically, SUMO-targeted ubiquitin ligases (STUbLs) have emerged as key players in nuclear quality control, genome maintenance, and transcription. However, how STUbLs select specific substrates among myriads of SUMOylated proteins on chromatin remains unclear. Here, we reveal a remarkable co-localization of the budding yeast STUbL Slx5/Slx8 and ubiquitin at seven genomic loci that we term "ubiquitin hotspots". Ubiquitylation at these sites depends on Slx5/Slx8 and protein turnover on the Cdc48 segregase. We identify the transcription factor-like Ymr111c/Euc1 to associate with these sites and to be a critical determinant of ubiquitylation. Euc1 specifically targets Slx5/Slx8 to ubiquitin hotspots via bipartite binding of Slx5 that involves the Slx5 SUMO-interacting motifs and an additional, novel substrate recognition domain. Interestingly, the Euc1-ubiquitin hotspot pathway acts redundantly with chromatin modifiers of the H2A.Z and Rpd3L pathways in specific stress responses. Thus, our data suggest that STUbL-dependent ubiquitin hotspots shape chromatin during stress adaptation.
染色质是一个高度调控的环境,蛋白质与染色质的结合通常受到翻译后修饰和相应的酶机制的控制。具体来说,SUMO 靶向泛素连接酶(STUbLs)已成为核质量控制、基因组维护和转录的关键因素。然而,STUbLs 如何在染色质上众多的 SUMO 化蛋白中选择特定的底物仍然不清楚。在这里,我们揭示了酿酒酵母 STUbL Slx5/Slx8 和泛素在我们称之为“泛素热点”的七个基因组位点上的显著共定位。这些位点的泛素化依赖于 Slx5/Slx8 和 Cdc48 分拣酶的蛋白周转。我们确定转录因子样 Ymr111c/Euc1 与这些位点相关,并成为泛素化的关键决定因素。Euc1 通过 Slx5 的二部分结合特异性靶向 Slx5/Slx8 到泛素热点,该结合涉及 Slx5 的 SUMO 相互作用基序和另外的新型底物识别结构域。有趣的是,Euc1-泛素热点途径在特定应激反应中与 H2A.Z 和 Rpd3L 途径的染色质修饰物冗余地作用。因此,我们的数据表明,STUbL 依赖性泛素热点在应激适应过程中塑造染色质。